����זE�̍R����܊������߂���@
�y�u����זE�̍R����܊������߂���@�v���K�v�Ȃ킯�z
�}�������a�∫�������p��̂悤�����t�����̏ꍇ�́A�R������ÂŎ���ꍇ������܂��B���t����ɑ���R������Â̐��������ƂɁA�Ō`�����ł��S�g�ɓ]�ڂ��Ď�p����ː����Â̓K���ɂȂ�Ȃ��ꍇ�̎��Ö@�Ƃ��čR������Â��s����悤�ɂȂ�܂����B
�������A���t����قǂ̌��ʂ��F�߂��Ȃ��Ƃ��������ɒ��ʂ��邱�ƂɂȂ�܂����B���̗��R�́A������Ō`����́A�}�������a�∫�������p��̂悤�Ȍ��t����Ƃ͈قȂ�l�X�ȗ��R�������āA�R����܂������ɂ����Ȃ��Ă��邩��ł��B
�Ⴆ�A�}�������a�∫�������p��͎�ᇍזE�̐������ψ�ŁA�זE����̕p�x�������̂ŁA����p���ς�����M���M���̍��p�ʂ̍R����܂𓊗^������A�����ڐA�p���鍂�p�ʂ̍R������Âɂ���āA���זE���܂߂Ď�ᇍזE��S�ł��邱�Ƃ��ł��܂��B
�������A�������āA���t�z���s�\���ȕ��ʂ�����i�R����܂����B���ɂ����j�Ō`����̏ꍇ�́A���Z�x�̍R������Â��s���Ă��A���זE�������c��\���������Ƃ����܂��B �܂��A����g�D���̂��_�������Ă���Ō`������_�f�̂���g�D�ɂ́A�R����܂������ɂ����Ƃ������R������܂��B
�ʏ�̍R������ẤA����p���ς�����ő�ʂ𓊗^���Ă���זE��Z���ԂŎ��ł�������@����{�ɂȂ��Ă��܂��B�R����܂ɑ���ϐ����o�Ă���O�ɒZ���Ԃɂ���זE��S�ł���������ǂ��ƍl���邩��ł��B
�}�������a�∫�������p��ł͂��̕��@���L���ł��B
�������A�Ō`����̏ꍇ�́A���̕��@�͕K�������L���ł͂���܂���B
������ᇂ⏬�זE���x����̂悤�ɍR������Â������������Ō`���������܂����A�����̌Ō`����i�x������X�������݂���Ȃǁj�ɑ��ẮA�R����܂̌����ڂ͌���I�ł��B
���̗��R�́A�O�q�̂悤�ȒZ������ɂ����Ă����Ă��A���t����ƈ���ČŌ`����͍R����܂������ɂ�������ł��B
�����A�R������Â͎�p����ː����Â��ł��Ȃ��}�������a�∫�������p��ɑ��ĊJ�����ꂽ���Ö@�ŁA��������̂܂܌Ō`����ɉ��p�����_�ɖ���������̂�������܂���B
�������A�Ȃ������Ȃ����Ƃ������R������A�������������A�R������Â��Ō`����ɂ��������ƂɂȂ�܂��B
�܂�A�u�Ō`����ɂ�����R����܊������߂��v���Ƃ��ł���A���Â̖��_�̑����������ł��邱�ƂɂȂ�܂��B
���Âɂ�������̑����́A�u�Ō`����ɑ���R������Â̗L�������Ⴂ�v���ƂƁu�R������Â̕���p�������v���Ƃ̂Q�_�ɋN�����Ă��āA������������邱�Ƃ��A����̕⊮�E��ֈ�Â̍ő�̖ړI�ɂȂ��Ă��܂��B
�u�R����܂͌����Ȃ��v�u����p�ŋꂵ�ށv�Ɣے�⋑�ۂ�������A�u�R����܂��������@�╛��p���y��������@�������āA�������H����v���Ƃ���茚�ݓI���Ǝv���܂��B
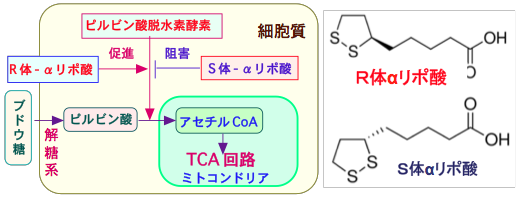
�y�R����܂𓊗^����Ƃ��̍R����܂ɑϐ��̍זE�������c��z
����זE�����ł�����R������Â͂���̑����h����v�Ȏ�i�ł��B �������A���p�ʂ̍R����ܓ��^�ɂ���čŏ��͎�ᇂ̑����}���Ă��A����ɍR����܂ɒ�R���̂���זE�������āA�R����܂ł���̑����}���邱�Ƃ��ł��Ȃ��Ȃ�܂��B
�Ⴆ�Δ������܂��V�X�v���`���͑����̂���̎��ÂɎg���Ă��܂��B �V�X�v���`����2�̉��f���q���ʂ�DNA�ƌ������邽�߁ADNA�����ɉˋ����`������ADNA������j�Q���A�זE���Ă��邪��זE����ѐ���זE�����ł����܂��B
�ŏ��͗ǂ������Ă��k������̂ł����A�R������Â��p�����Ă���Ǝ���ɃV�X�v���`���ɒ�R���̂���זE�������Ă����܂��B
�R����܂��n�߂�O����A���̂���g�D�ɂ̓V�X�v���`���Ɋ��̍זE�ƒ�R���̍זE�����݂��A�V�X�v���`���̓��^�ɂ���āA�V�X�v���`�����̂���זE�����ł��A�V�X�v���`����R���̂���זE�������c��A����ɃV�X�v���`����R���̂���זE�������邽�߂ɁA����ɍR����܂������Ȃ��Ȃ�܂��B�i�}�P�j
|
�}�P�F�R������ÑO�̂���g�D�̂���זE�͖�ܑϐ��̒��x�ɂ����ĕs�ψ�ŁA��ܑϐ��̒Ⴂ����זE�⍂������זE�����݂��Ă���i�@�j�B�R����܂̃V�X�v���`�����^���s���ƁA�V�X�v���`���Ɋ��̂���זE�͎��ł��邪�A�ϐ��̂���זE�͐����c��i�A�j�B�����c��������זE�͑��B���Ă���͍Ĕ�����i�B�j�B����ɃV�X�v���`���𓊗^���Ă��A����זE�͎��ł����ɂ���ɃV�X�v���`���ϐ��זE�̔䗦�͑����A����g�D�͑��傷��i�C�j�B |
�V�X�v���`�����^���V�X�v���`���ϐ�����זE�𑝂₷�Ƃ����̂��_�[�E�B���̐i���_�Ɠ����ł��B�V�X�v���`���Ŏ��ł���זE�͎��R��������Ďq�����c�����ɖłсA�V�X�v���`���ɒ�R���̂���זE�������I��I�ɐ����c��܂��B �R������Â�����זE�̐i���̑I�����Ƃ��ē����܂��B
�R������Â��n�܂�A����זE�ԂŐ����������J�n����܂��B �R����܂��g�p����ƍR����܂ɒ�R���̍זE���c�点��Ƃ��������I�����������܂��B �ʏ�̍R������ẤA��ܑϐ��̏o���Ɛ����𑣐i����Ƃ������_�������Ă��܂��B
�y����זE�͗l�X�ȃ��J�j�Y���ōR����ܑϐ����l������z
�����ł́A�u����זE�̍R����܊������߂���@�v�͏d�v�ȗ̈�ł���A��b������Տ������ő����̌������ʂ�����Ă��܂��B �u�R����܊������߂�v�Ƃ����͍̂R����܂̌����ڂ����߂���@�ł��B
�P���ɍR������Â��������{���Ă��A���̌��ʂɂ͌��E������܂��B��ʓI�ɁA�Ō`����ɑ��Ă͍R����܂����ł͂���זE�����łł��܂���B
����ɁA�R����܂��g�������Ă���ƁA���������Ȃ��Ȃ�Ƃ������܂��B����זE�����̍R����܂ɑϐ����l�����邽�߂ł��B
�Ⴆ�Δx�B����ŕ��q�W�I��̃`���V���L�i�[�[�j�Q�܁i�C���b�T�A�^���Z�o�Ȃǁj�������Ă��A������10����14�������x�Ō����Ȃ��Ȃ�܂��B �ʏ�̎E�זE��p���������R������Â��A����������P�N������Ό����Ȃ��Ȃ��Ă��܂��B
����זE���R����܂Ń_���[�W����ƁA�����ȃ��J�j�Y�����g���Đ����c���i���l�����Ă��܂��B ���ÑO���炪��g�D�̒��ɑϐ��זE�����݂����������iintrinsic�j�̌��������łȂ��A�R������Â���ܑϐ��̐����i�����l�����iacquired�j�̍R����ܑϐ�������܂��B
�R����܂��זE�̊O�ɔr�o����|���v�̍�p�����`�����̍�������������A�R����܂�s���������镨�����̖ڕW�ɂȂ�`�����Y�����Ė�܂̍�p��W�Q���܂��B�R����܂̍U���ڕW��DNA�ł���ADNA�C���𑣐i���邱�Ƃɂ��זE����j�~���悤�Ƃ��܂��B
�����̍R����܂͍זE�̃A�|�g�[�V�X�������N�������Ƃɂ���Č��ʂ����܂��B�זE�ɂ͎���זE�������s����v���O���������݂��Ă���A�זE�����t���Ƃ��̃v���O�����ɂ���Ď��ɂ܂��B���̍זE�����A�|�g�[�V�X�Ƃ����܂��B
����זE�͂����ȋ@���ɂ���ăA�|�g�[�V�X�ɑ����R�����l�����Ă����܂��B�זE�����N���ɂ����Ȃ�Ƃ����A�|�g−�V�X�ϐ��̊l�����R����ܑϐ��̏d�v�ȃ��J�j�Y���ł��i�}�Q�j�B
|
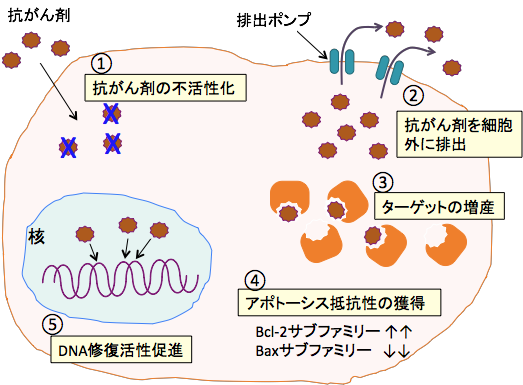
�}�Q�F����זE�͗l�X�ȃ��J�j�Y���ōR����܂̌����ڂ���߂Ă���B�Ⴆ�A�R����܂̕������ӂɂ��s�������̑��i�i�@�j�A�r�o�|���v�𑝂₵�čR����܂��זE�O�ւ̔r�o�̑��i�i�A�j�A�R����܂̃^�[�Q�b�g���q�̑��Y�i�B�j�A�A�|�g�[�V�X�ɒ�R���ɂȂ�Bcl-2�T�u�t�@�~���[�̃^���p�N���𑝂₵����A�A�|�g�[�V�X��U������Bax�T�u�t�@�~���[�̊�����}�����čזE���ɑ����R���̊l���i�C�j�A�_���[�W����DNA�ȂǍזE�����̏C���̑��i�i�D�j�A�ȂǑ����̃��J�j�Y�����m���Ă���B |
�y�R����ܑϐ��ɂȂ�ɂ̓G�l���M�[���K�v�z
��ʂɁA�R����ܒ�R���̂���זE�͑��B���x�����Ƃ��m���Ă��܂��B�R����܂ɒ�R���̋@��������̂ɗ]���ȃG�l���M�[�ƕ����������K�v������ł��B
�Ⴆ�A�R�_���������߂邱�Ƃɂ���čR����ܒ�R���ɂȂ�܂����A���̍R�_���V�X�e�������߂�ɂ͗]�����G�l���M�[�iATP�j���������������i�O���^�`�I����R�_���y�f�Ȃǂ̎Y���j���K�v�ł��B
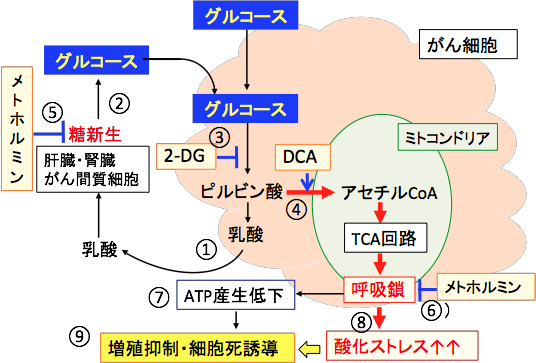
�����������A�R����܊������߂���@�̑��́A����זE�̃G�l���M�[�Y����ቺ�����邱�Ƃł��B
����זE�͉n�̑�ӂ����i���Ă��܂��B�n��j�Q����2-�f�I�L�V-D-�O���R�[�X�ƃ~�g�R���h���A��ATP�Y����j�Q�������g�z���~���̕��p�͂���זE�̃G�l���M�[�Y����j�~���āA�R������Â���ː����Â̌����ڂ����߂܂��B
�y���g�z���~���̓~�g�R���h���A�̌ċz�y�f������I��j�Q����z
���g�z���~���imetformin�j�́A���E���łP���l�ȏ���Q�^���A�a�����Ɏg���Ă���r�O�A�i�C�h�n�o�������~���܂ł��B �r�O�A�i�C�h�܂́A�������Y�̃}���Ȃ��K���K�iGalega officinalis�j����1920�N��Ɍ��������O�A�j�W���U���̂���J�����ꂽ��ł��B�K���K�͌Â�����A���A�a�Ǝv����a�C�i�����⑽�A�j�̎��ÂɌo���I�Ɏg���L�������F�߂��Ă���A���̊W�ł��̃K���K���猌���~����p�̂���r�O�A�i�C�h����������܂����B
���g�z���~���́A�~�g�R���h���A�̌ċz���̍ŏ��̃X�e�b�v�ł����ċz�y�f������I��j�Q���邱�Ƃ����炩�ɂȂ��Ă��܂��B���̌��ʁA�~�g�R���h���A�ł�ATP�Y�����������AAMP:ATP�̔䂪�㏸���AAMP�������v���e�C���L�i�[�[�iAMPK�j������������܂��B
����������AMPK�́A�̑��̓��V����}�����A�i���A���i�ł̃O���R�[�X���p�𑣐i���Č�����ቺ�����܂��BAMPK�̓C���X�����������߂�̂ŁA���Ȃ��C���X�����Ō������R���g���[���ł���悤�ɂȂ�܂��B
�C���X�����͘V���Ɣ�����𑣐i���A����זE�̑��B�𑣐i����̂ŁA���A�a�łȂ��l�ł��R�V���Ƃ���\�h�̖ړI�ŕ��p���Ă���l�����܂��B
���Ȃ킿�A���g�z���~���̌����~����p�̓~�g�R���h���A�ɂ�����ATP�Y���̑j�Q�ɂ���đ̓���ATP�ʂ��������邽�߂ł��B�̂�ATP�𑝂₷���߂ɁA�O���R�[�X�̕����i�ى��j�𑣐i���A���V���i�����j��}������̂ŁA�������ቺ���܂��B�^����ATP����������AMPK�������������̂Ɠ������J�j�Y���ł��BATP�Y������������̂�AMPK�����������܂��B
���̃��g�z���~���̌ċz�y�f�j�Q��p�́A����זE�ɂ����Ċ����_�f�̎Y���𑝂₷�ړI�ł��Âւ̉��p����������Ă��܂��B
|
|
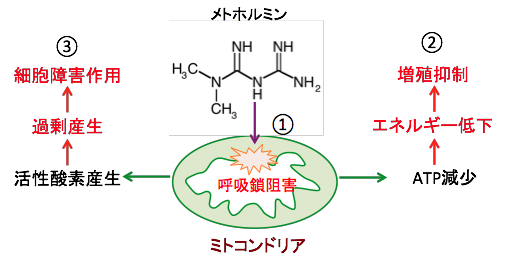
�}�R�F���g�z���~���̓~�g�R���h���A�̌ċz�y�f�����̂P��j�Q����i�@�j�B���̌��ʁAATP�Y�����������ăG�l���M�[�ቺ�ɂ���đ��B��}�������i�A�j�B�ċz���̑j�Q�ɂ���ă~�g�R���h���A�ł̊����_�f�̎Y���������A�_���X�g���X�ōזE�͏�Q�����i�B�j�B |
�y2-�f�I�L�V-D-�O���R�[�X�͉n��j�Q����z
2-�f�I�L�V-D-�O���R�[�X�i2-Deoxy-D-glucose�j�́A�O���R�[�X�i�u�h�E���j��2�ʂ̐��_��iOH�j�����f���q�iH�j�ɒu�����ꂽ�����i�O���R�[�X�U���́j�ł��B 2-�f�I�L�V�O���R�[�X(2-DG)�̓O���R�[�X�Ɠ����悤�ɃO���R�[�X�A���́i�O���R�[�X�E�g�����X�|�[�^�[�j��GLUT1�𗘗p���čזE���Ɏ�荞�܂�܂��B
�O���R�[�X��2-DG�͍זE���ɓ���ƃw�L�\�L�i�[�[�ɂ���ă����_������A�O���R�[�X-6-�����_���邢��2-�f�I�L�V-D-�O���R�[�X-6-�����_�i2-DG-6-�����_�j�ɕϊ�����܂��B�����_�������ƃO���R�[�X�E�g�����X�|�[�^�[��ʉ߂ł��Ȃ����ߍזE�O�֏o��Ȃ��Ȃ�܂��B
���̃w�L�\�L�i�[�[�ɂ��U�ʂ̃����_���͉n�ɂ��u�h�E���i�O���R�[�X�j�̑�ӂ̍ŏ��̃X�e�b�v�ŁA�זE���Ɏ捞�u�h�E�����זE���ɂƂǂ߂Ă����ړI������܂��B�����_��������́A�O���R�[�X-6-�����_�͂�����n�ő�ӂ���ăG�l���M�[�Y���Ɏg���A�y���g�[�X�E�����_�o�H�Ŋj�_�Ȃǂ̕��������̍ޗ��Ƃ��Ă����p����܂��B
�������A2-DG-6-�����_�́A�n�y�f�ő�ӂł��Ȃ����߁A�זE���ɒ~�ς��܂��B�O���R�[�X-6-�����_��2-DG-6-�����_��E�����_������t�H�X�t�@�^�[�[�����V�����s���̑���t���̍זE�ɂ͂���܂����A�����̂���זE�̓t�H�X�t�@�^�[�[�̊������Ⴂ�̂ŁA��U������2-DG��2-DG-6-�����_�ɕϊ����ꂽ���Ƃ͍זE�O�ɏo�邱�Ƃ��ł����A����ɂ���ȏ��ӂ���邱�Ƃ��ł��Ȃ��̂ŁA2-DG-6-�����_�̏�Ԃłǂ�ǂ�~�ς��܂��B
2-DG�ɂ���ăG�l���M�[�Y�����ቺ����Ƃ��̃X�g���X�����ɂ���ăO���R�[�X�g�����X�|�[�^�[�̔���������ɑ����A2-DG�̎�荞�݂�����ɑ��₷���Ƃɂ��Ȃ�܂��B���������āA����זE�͐���זE�ɔ�ׂĂ��2-DG�̎捞�݂������܂��B
�זE���Œ~�ς���2-DG-6-�����_�̓w�L�\�L�i�[�[�ƃw�L�\�[�X�E�t�H�X�t�F�[�g�E�C�\�����[�[��j�Q���܂��i�h�R�j�Q�j�B���������āA2-DG���o���ێ悷��ƁA����זE�ɑ�����荞�܂�A����זE�̉n��j�Q����̂ŁA�O���R�[�X�i�u�h�E���j�̑�ӂɂ��G�l���M�[�Y���ƕ���������j�Q���邱�ƂɂȂ�܂��B
2-DG������זE���ɑ����g���b�v����邱�Ƃ𗘗p���������@���o�d�s�ł��B�o�d�s�́u�|�W�g�����E�G�~�b�V�����E�g���O���t�B�[�iPositron Emission Tomography�j�v�̗��ŁA���{��ł��z�d�q���ː��f�w�B�e�Ƃ����܂��B
2-DG�̂Q�ʂ̐��f���q�i�܂�A�O���R�[�X�̂Q�ʂ�OH��j��z�d�q���o���ʑ̃t�b�f18�i18F�j�Œu�����ꂽ18F-�t���I���f�I�L�V�O���R�[�X�iFDG�j�Ƃ�����܂𒍎˂�����A������o�d�s���u�ŎB�e���AFDG�̏W�܂�����摜�����Đf�f������̂ł��B
�����̂���́A�O���R�[�X��荞�݂���уw�L�\�L�i�[�[���x�����㏸���Ă��邽�߁A����זE��FDG���W�܂�̂ł��B 2-DG�͂���זE�ɗD��I�Ɏ�荞�܂�A�n��y���g�[�X�E�����_�o�H��j�Q���āA����זE�������������邱�Ƃ��ł���̂ł��B
|
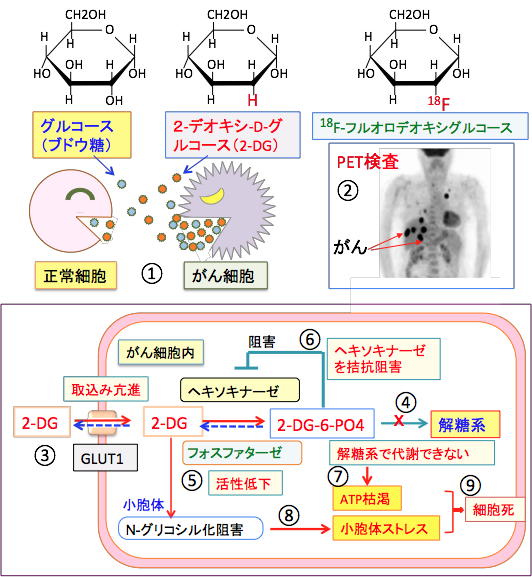
�}�S�F2-�f�I�L�V-D-�O���R�[�X�i2-Deoxy-D-glucose�j�́A�O���R�[�X�i�u�h�E���j��2�ʂ̐��_��iOH�j�����f���q�iH�j�ɒu�����ꂽ�����i�O���R�[�X�U���́j�ŁA����זE�̓O���R�[�X�̎捞�݂������A2-DG�̎捞�݂������i�@�j�B2-DG������זE���ɑ�����荞�܂�邱�Ƃ𗘗p���������@���o�d�s�iPositron Emission Tomography�j�i�A�j�B2-�f�I�L�V-D-�O���R�[�X�i2-DG�j�̓O���R�[�X�E�g�����X�|�[�^�[�iGLUT1�j�ɂ���čזE���Ɏ�荞�܂��i�B�j�B����זE��GLUT1�̔����ʂ������A�O���R�[�X�Ɠ�����2-DG��������荞�ށB2-DG�̓w�L�\�L�i�[�[��2-DG-6-�����_�i2-DG-6-PO4�j�ɕϊ�����邪�A����ȏ��ӂ���Ȃ��i�C�j�B����זE�̓t�H�X�t�@�^�[�[�̊������Ⴂ�̂ŁA2-DG-6-�����_������זE���ɒ~�ς���i�D�j�B2-DG-6-�����_�̓w�L�\�L�i�[�[���t�B�[�h�o�b�N�j�Q�i�A���X�e���b�N�j�Q�j����̂ŁA 2-DG-6-�����_����荞����זE�̓O���R�[�X�̉n�ł̑�ӂ��j�Q�����i�E�j�B���̌��ʁA����זE�̃G�l���M�[�Y���ƕ��������͑j�Q����邱�ƂɂȂ�AATP���͊�����i�F�j�B2-DG�͏��E�̂ł̃^���p�N����N-�O���R�V�����i�����̌����ɂ��C���j��j�Q���A�܂��݂̕s���S�Ȉُ�^���p�N���iunfolded protein�j�𑝂₵�ď��E�̃X�g���X�������N�����i�G�j�B���̌��ʁA����זE�͎��ł��₷���Ȃ�A�R����܊������i����i�H�j�B |
2-DG������זE�̑��B��}��������ʂ��w�E���ꂽ�̂�1950�N��ł��B�u�זE�̃G�l���M�[���ł���O���R�[�X�̗U���̂���荞�܂���A����זE�̑��B��}���ł����v�Ƃ����A�C�f�A�́A����70�N���O�Ɍ�������Ă���A�O���R�[�X�̗U���̂̍R��ᇊ�������������A2-DG�ɋ����R��ᇌ��ʂ����邱�Ƃ��ؖ�����Ă��܂��B
�������A2-DG���g�������ẤA���̌゠�܂蒍�ڂ���Ȃ������悤�ł��B���̗��R�̈�́A����̎��Âɂ����ẮA�u�����Ő������������������g���Ă���זE����|����悤�Ȏ��Ö@�v��1950�N��ȍ~�͎嗬�ɂȂ��Ă������炾�Ǝv���܂��B���̂��߁A�u�G�l���M�[�Y���o�H��j�Q���Ă���זE�̑��B��ቺ������v�Ƃ����悤�ȃA�C�f�A�͒��ڂ���Ȃ������̂�������܂���B
�������A���[���u���O���ʂ��ĕ]�������悤�ɂȂ�A����זE�̃G�l���M�[�Y���ƕ���������j�Q������@�Ƃ��āA2-DG�ɂ����ڂ��W�܂�悤�ɂȂ�A�����̓��������ōR��ᇌ��ʂ��ؖ�����A�l�Ԃł̗Տ����������{�����悤�ɂȂ����Ƃ������Ƃł��B
�y2-�f�I�L�V-D-�O���R�[�X�͍R������Â���ː����Â̌����ڂ����߂�z
�Q-�f�I�L�V-D-�O���R�[�X�i2-DG�j�͂���זE�̉n��j�Q����̂ŁA����זE�̑��B���x��ቺ��������ʂ�����܂����A�Q-DG�P�Ƃł͂���זE�����ł������p�͎ア�ƌ��킴������܂���B���܂ŁA����������l�Ԃł̌���������Ă��܂����A2-DG�̓��^�����ł͏\���ȍR��ᇌ��ʂ͓����Ă��܂���B����זE�̃u�h�E�������S�Ɍ͊������邱�Ƃ������I�ɍ������ł��B
�@
�������A����זE�̃G�l���M�[�Y���╨�������̌o�H��j�Q����ƁA�R����܂���ː��ɑ��邪��זE�̊������܂�܂��B�R������Â���ː����Â̎��ɂQ-DG�p����ƁA�����̍R��ᇌ��ʂ����߂邱�Ƃ������̗Տ������Ŋm�F����Ă��܂��B �@
�R����܂Ƃ̕��p�ɂ������P���̏d�Pkg������40�`60mg���x�̓��^�ʂŗՏ��������s���Ă��܂��B�������^�̈��S���͏\���Ɍ�������Ă��Ȃ����߁A����̍Ĕ��\�h�̖ړI�ł͂܂������ł��܂��A�i�s����̎��Â̖ړI�ōR����܂ȂǂƂ̕��p�ȂǒZ���Ԃ̎g�p�Ɋւ��Ă͖�薳���悤�ł��B
�@
2DG�ƃO���R�[�X�i�u�h�E���j���������Ă���זE�̃G�l���M�[��ӂ�j�Q���邽�߁A���������ŃO���R�[�X�̐ێ�ʂ����点�A2DG�͏��Ȃ��ʂőj�Q��p���ł��܂��B �@
�A�Q���͋ؓ���S����]�̓������ቺ���Č�����O���R�[�X�̎捞�݂�����܂��B���̂��߁A2-DG���A�Q�O�ɕ��p����ƁA�ł��R��ᇌ��ʂ����܂�܂��B �@
2-DG�̓Ő��Ɋւ��ẮA�}�E�X�̎����ł�50%�v���ʂ�2g/kg�ȏ�Ƃ���������܂��B�iCent Eur J Biol. 5:739–748. 2010�N�j �l�ł̌����ł�200mg/kg���炢�܂ł͓��^�ł���Ƃ���������܂��B
�@
�ł���������p�͍������ł��B2-DG�͍זE���̃O���R�[�X�̔Z�x��ቺ�����܂��B�]�̎��������̐_�o�זE���זE���O���R�[�X�̒ቺ�����m����ƁA�ጌ����ԂƊ��Ⴂ���āA�]�����̂̃z����������𐧌䂵�Č��������߂�z��������`�B�������o���悤�ɂȂ邽�ߍ������ɂȂ�悤�ł��B�H������̓����ێ�����炷�P�g���H��̑��̓��V����j�Q���郁�g�z���~���p����ƍ�����������邱�Ƃ��ł��܂��B
����A���p�ʂ������ƒጌ���̂悤�ȏǏ�i���ӊ���E�́j�������܂��B����זE�ɑ����捞�܂�邽�߁A�ጌ���ǏN����Ȃ����x���ŕ��p�ʂ߂��邱�Ƃ��d�v�ł��B
�y2-DG�͐���זE���R����܂���ː��̃_���[�W����ی삷���z
�Q-�f�I�L�V-D-�O���R�[�X�i2-DG�j�͂���זE�̉n��j�Q����̂ŁA����זE�̑��B���x��ቺ��������ʂ�����܂��B ����זE�̃G�l���M�[�Y���╨�������̌o�H��j�Q����ƁA�R����܂���ː��ɑ��邪��זE�̊������܂�܂��B
�Q-�f�I�L�V-D-�O���R�[�X�i2-DG�j�͍R����܂���ː��ɑ��邪��זE�̊������߂邾���łȂ��A�R����܂���ː��ɂ�鐳��זE�̃_���[�W���y��������ʂ�����Ƃ���������܂��B�ȉ��̂悤�ȕ�����܂��B
Protection of normal cells and tissues during radio- and chemosensitization of tumors by 2-deoxy-D-glucose.�@(2-�f�I�L�V-D-�O���R�[�X�͂���g�D�̕��ː����ƍR����܊������߁A����זE�Ƒg�D�̃_���[�W���y������)J Cancer Res Ther. 2009 Sep;5 Suppl 1:S32-5.
����זE�͐���זE�ɔ�ׂăO���R�[�X�i�u�h�E���j�̎捞�݂������AATP�Y����זE���邽�߂̕��������ɑ�ʂ̃O���R�[�X��K�v�Ƃ��Ă��܂��B���������āA�O���R�[�X�̎捞�݂◘�p��W����AATP�Y���╨���������ቺ���A�R����܂���ː����Â̌����ڂ������Ȃ�܂��B
����זE�̓O���R�[�X�g�����X�|�[�^�[�𑽂��������Ă���̂ŁA2-DG�̎捞�݂������A2-DG�ɂ��O���R�[�X��ӂ̑j�Q��p�������Ɍ���܂��B
�|�{�זE���g���������⓮���ɂ���זE���ڐA�������������ŁA2-DG�𓊗^����ƍR����܂���ː����Â̎��Ì��ʂ����܂邱�Ƃ������̎����n�Ŋm�F����Ă��܂��B
����ɓ��������ŁA2-DG���]��S���ɑ���R����܂���ː��̃_���[�W���y�������p���F�߂��Ă��܂��B���̍�p�@���ɂ��Ă͂܂��\���ɉ𖾂���Ă��܂��AAMP�������v���e�C���L�i�[�[�iAMPK�j�̊�������I�[�g�t�@�W�[�̑j�Q�ȂǕ����̃��J�j�Y������������Ă��܂��B
�ȉ��̂悤�Ș_��������܂��B
Caloric restriction mimetic 2-deoxyglucose antagonizes doxorubicin-induced cardiomyocyte death by multiple mechanisms.�i�J�����[�����Ɠ��l�̍�p������2-�f�I�L�V�O���R�[�X�̓h�L�\���r�V���ɂ��S�؍זE�����̃��J�j�Y���őj�~����jJ Biol Chem. 2011 Jun 24;286(25):21993-2006.
�H������̃J�����[�ێ�����炷�J�����[�������S���njn�̌��N��Ԃ�ǂ����邱�Ƃ��m���Ă��܂��B�O���R�[�X�މ�������2-�f�I�L�V-D-�O���R�[�X�̓J�����[�����Ɠ��l�̍�p���������Ƃ������̓��������ŕ���Ă��܂��B
���̘_���ł́A�R����܂ŕ���p�Ƃ��ĐS�؏�Q�������N�����h�L�\���r�V���̓��^�ň����N�������S�؍זE���ɑ���2-DG���}����p���������ǂ������������Ă��܂��B�����̌��ʁA2-DG�̓h�L�\���r�V���ŗU�������S�؍זE�̃_���[�W��זE����j�~���邱�Ƃ�������Ă��܂��B
2-DG�͗l�X�ȃ��J�j�Y���ōR��ᇍ�p�������A���ɍR����܂���ː����ÂƂ̕��p�ŁA�R��ᇌ��ʂ����߂邾���łȂ��A����זE��ی삷���p������̂ŁA���Â̕⊮�Ƃ��ė��p���l�͍����ƌ����܂��B
�y����זE�̎_���X�g���X�����߂�Ǝ��ł��₷���Ȃ�z
����זE�ł͎_�f���\���ɑ��݂���ꍇ�ł��A�n�ł̃O���R�[�X��ӂ����i���āA���ΓI�Ƀ~�g�R���h���A�ł̎_���I�����_���͗}������Ă��܂��B���̌��ʁA���_�̎Y���������Ă��܂��B������D�C�I�����邢�����[���u���O�����ƌ����܂��B
�~�g�R���h���A�ł̎_���I�����_����}���Ă���̂́A�����_�f�̊Q�����炵��������ł��B �������A����ł��~�g�R���h���A�ł̎_���I�����_���ɂ��G�l���M�[�Y���͋N�����Ă��܂��B��ʓI�ɁA���B�̑�������זE�ł́AATP�Y����50%�ȏオ�n�ŎY�������ƌ����Ă��܂��B
����זE�̓~�g�R���h���A�@�\�ɂ����Ȉُ킪����̂ŁA����זE�ɔ�ׂĊ����_�f���������₷���Ȃ��Ă��܂��B�����ŁA����זE�͎_���X�g���X���y�����邽�߂ɁA�~�g�R���h���A�ł̎_���I�����_����}���A�זE�ɔ�������R�_���V�X�e���i���Ă��܂��B
�������R��������ɂ���Ĉ����N�������זE���i�A�|�g�[�V�X�j�́A�S�Ăł͂Ȃ��ɂ��Ă����̂قƂ�ǂ͊����_�f��ɂ���Ĉ����N�������\����������Ă��܂��B���ː��Ǝ��������_�f��̎Y���ɂ���ăA�|�g�[�V�X��U������̂Ɠ����悤�ɁA�����̍R����܂��ŏI�I�ɂ͊����_�f����Y�����邱�Ƃɂ���ăA�|�g�[�V�X��U�����Ă���̂ł��B
�����̍R����܂�����זE�ɃA�|�g�[�V�X��U�����鋤�ʂ̃��J�j�Y���Ƃ��Ċ����_�f���g���Ă���Ƃ������Ƃ́A�Ȃ��R����܂ɒ�R���̂���זE�͕��ː����Â����l�ɒ�R���ɂȂ邩�Ƃ������R��������Ă��܂��B
���̂悤�Ȋ����_�f����Y�������Ă���זE�����ł�����Ƃ��ɍR�_���܂�N-�A�Z�`���V�X�e�C����O���^�`�I�����ɓ��^����ƁA�����_�f�ɂ�邪��זE�̎��ł͋N����Ȃ��Ȃ�܂��B
���������āA�R����܂���ː����Â̌����ڂ����߂���@�Ƃ��āA�~�g�R���h���A�ł̊����_�f�̗ʂ𑝂₷���@�i�ċz�y�f������-I�̑j�Q�{�_���I�����_���̊������j�ƍR�_���V�X�e���i�O���^�`�I���A�`�I���h�L�V���Ȃǁj��j�Q������@��g�ݍ����邱�Ƃ��L�p�ł��邱�Ƃ������ł��܂��B
�y�d�q�`�B�n�i�ċz���j����̘R�ꂪ�����_�f��̗ʂ����߂Ă���z
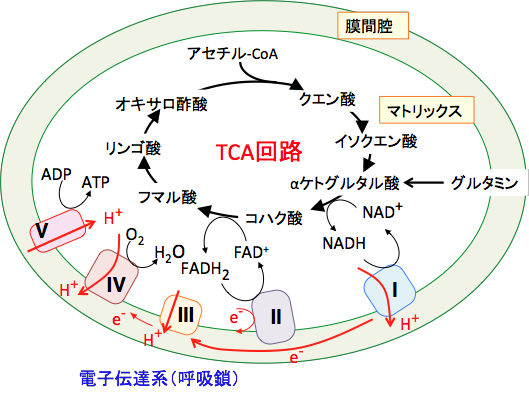
�~�g�R���h���A�ɂ������d�q�`�B�n�i�ċz���j�ɂ�����ATP���Y�������Ƃ��A�K�R�I�������_�f��i�X�[�p�[�I�L�V�h�A�ߎ_�����f�A�q�h���L�V���W�J���Ȃǁj���������܂��B �~�g�R���h���A��TCA��H�ɂ��NADH��FADH2�̌`�ŕߑ����ꂽ���f�́C�~�g�R���h���A�ɂ����āC��A�̍y�f�n�i�ċz�������� I�`IV�j��ATP�����y�f�i�ċz��������V�Ƃ������j�̘A�����o�āC�ŏI��e�̂ł���_�f�iO2�j�ɓn����Đ��iH2O�j�ɂȂ�܂��B
������ I�`IV�̒i�K�́C�~�g�R���h���A�����̃^���p�N�����y�f�Ԃœd�q�̂���肪�N����ߒ��ł��邽���d�q�`�B�n�i�ċz���j�ƌĂ�܂��B �d�q�`�B�n�ɂ���ă~�g�R���h���A�}�g���b�N�X���疌�ԋ�Ԃ��v���g���i���f�C�I���j�����ݏo����A�A�����ꂽ�v���g���ɂ���ă~�g�R���h���A�����̓��O��ΔΨ�ƌĂ���d�C���w�I�|�e���V�����i�v���g���ɂ���Đ�����d�ׂ̍��j�����o����܂��B
�}�g���b�N�X���ɖ߂�v���g���̋쓮�͂𗘗p����ATP�����y�f��ADP�Ɩ��@�����_����ATP���������܂��B������_���I�����_���ƌ����܂��B
�~�g�R���h���A�̌ċz����_���I�����_���̉ߒ����j�Q�����ƁA�v���g���i���f�C�I���j���������āA�~�g�R���h���A����̊����_�f��̎Y�����������܂��B���g�z���~���͌ċz�y�f������I��j�Q���āA�~�g�R���h���A����̊����_�f�̔����𑝂₷��p������Ă��܂��B
|
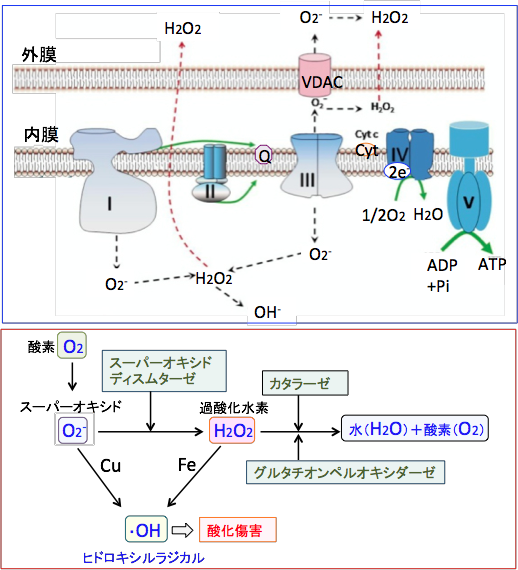
| �}�T�F�d�q�`�B�̍ۂɁA�ċz�y�f������I�╡����III����R��o�����d�q�ɂ���āA�_�f���q����d�q�Ҍ�����A�X�[�p�[�I�L�V�h�iO2-�j����������B�~�g�R���h���A�ŏ�����_�f��1�`3%�������_�f��ɕϊ������Ɛ�������Ă���B |
|
�}�U�F�ċz�y�f�����̂�I��III�ŃX�[�p�[�I�L�V�h�iO2-�j���Y�������B�X�[�p�[�I�L�V�h�̓X�[�p�[�I�L�V�h�E�f�B�X���^�[�[�iSOD�j�ɂ���ĉߎ_�����f�iH2O2�j�ɕϊ������B�ߎ_�����f�̈ꕔ��Fenton/ Haber-Weiss�����ɂ���ăq�h���L�V�����W�J���iHydroxyl radical�j�ɕϊ������B |
�y����זE�ł̓s���r���_�E���f�y�f�L�i�[�[�̊��������i���Ă����z
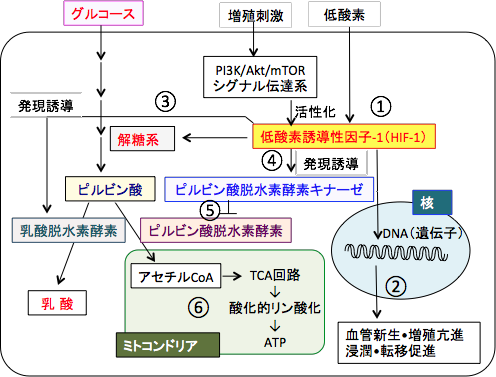
����זE�̑�ӂ̓����ł���u�n�̘��i�ƃ~�g�R���h���A�ł̎_���I�����_���̗}���v�Ƃ����A���������[���u���O���������{�Ő��䂵�Ă���̂���_�f�U�����q-1�iHypoxia-inducible Factor-1�FHIF-1�j�Ƃ����]�ʈ��q�ł��B �@
�]�ʈ��q�Ƃ����͓̂���̈�`�q�̔����iDNA�̏������b�Z���W���[RNA�ɕϊ����邱�Ɓj�߂��Ă���`�����ł��BHIF-1�̃^�[�Q�b�g��`�q��100��ވȏ�m���Ă���A�G�l���M�[��ӁA���ǐV���A�זE���B�A�A�|�g�[�V�X�ȂǍזE�̋@�\�Ɛ[���֘A���Ă����`�q�̔����𐧌䂵�Ă��܂��B �@
HIF-1�͍זE����_�f��Ԃɂ������Ɗ��������Ă��܂��B���������āA�_�f���\���ɗ��p�ł���ōזE���Ă��鐳��זE�ł͕K�v���Ȃ��]�ʈ��q�ł��B
�@
����A�����̂���זE�ł́A��_�f��Ԃł����Ă��Ȃ��Ă�HIF-1�̊��������i���Ă��܂��B�}���ɑ��傷�邪��g�D�̒��ŁA����זE�͏�ɒ�_�f���h�{�ɂ��זE���̊댯�ɂ��炳��Ă��܂��B�����ŁA��_�f���h�{�ɂ��זE�����N�����Ȃ��悤�ɂ��郁�J�j�Y���Ƃ��Ă���זE��HIF-1���������߂Ă��܂��B
����́AHIF-1���������i���Ă���قǁA����זE�͒�_�f���h�{�Ő����ł���i���ɂɂ����j�Ƃ������Ƃ��Ӗ����Ă��܂��B �@
����זE�ł́A��`�q�ψقȂǂɂ���đ��B�̃V�O�i���`�B�n���P��I�ɘ��i���Ă���A���̌��ʂƂ���HIF-1�̊������P��I�ɘ��i���Ă��܂��B
�܂�A�_�f�������Ă��A����������_�f�̂悤�ȑ�ӂ��s���Ă���킯�ł��B
�s���r���_�E���f�y�f�L�i�[�[���s���r���_�E���f�y�f(�s���r���_����A�Z�`��CoA�ւ̕ϊ�����y�f)�������_�����Ċ�����ቺ�����܂��BHIF-1�̓s���r���_�E���f�y�f�L�i�[�[�̔����𑣐i���A����Ƀs���r���_������_�ւ̌��C���n�ɓ������_�E���f�y�f�iLDH�j�̔����𑣐i�����p������܂��B
�܂�AHIF-1�̓s���r���_����A�Z�`��CoA�ւ̕ϊ���j�Q����TCA��H�Ǝ_���I�����_���ł̑�ӂ�}�����A���C���n(�s���r���_������_�̕ϊ�)�i���܂��B�n�̓r���̃u�h�E���̑�ӎY������j�_�⎉�b�_��A�~�m�_�̍����𑣐i�����p������܂��B �@
����זE�ł�HIF-1�͒�_�f�ɂȂ����Ƃ���������������܂��A����זE�ł͑��B�V�O�i���ُ̈�Ȃǂɂ����HIF-1�͍P��I�Ɋ��������A�_�f������ł��_�f��������ԂƓ�����ӂ��s�����߁A����זE�ł͉n�����i���A�~�g�R���h���A�ł̎_���I�����_�����}������Ă��܂��B
�@
����זE�Ŋ���������Ă���HIF-1�́A����זE�ɂ��������_�E���f�y�f�̎Y�������߂܂��B�܂��AHIF-1�̓s���r���_�E���f�y�f�̊�����j�Q�����s���r���_�E���f�y�f�L�i�[�[�̈�`�q�����𑣐i���܂��B
���������āA����זE�ł́AHIF-1�̔������i�ɂ���āATCA��H�ւ����o�H���Ւf����A���C���n�̘��i�ɂ���ē��_����ʂɎY��������ӂ����i���Ă��܂��B �@
���G�ȃ��J�j�Y���ł����AHIF-1�̊������i�͂���זE�̑�ӈُ�̒��S�ɂȂ��Ă���̂ŁA��ӂ��^�[�Q�b�g�ɂ������Ö@�̗����ɖ𗧂��܂��B�i�}�V�j
|
�}�V�F��_�f��B�V�O�i���`�B�n�iPI3K/Akt/mTOR�j�̊������i�ɂ���Ē�_�f�U�����q-1�iHIF-1�j�̔����Ɗ��������i����i�@�j�BHIF-1�͍זE�j��DNA�ɍ�p���āA����זE�̑��B��]�ڂ𑣐i����l�X�Ȉ�`�q�̔����i����i�A�j�BHIF-1�͉n�y�f�i�w�L�\�L�i�[�[�Ȃǁj����_�E���f�y�f�̔����i���ĉn�̑�ӂi����i�B�j�BHIF-1�̓s���r���_�E���f�y�f�L�i�[�[�̔�����U������i�C�j�B���̃L�i�[�[�̓s���r���_�E���f�y�f��j�Q����i�D�j�B���������āAHIF-1�̊������i�ɂ���ă~�g�R���h���A�ł̎_���I�����_�����}�������i�E�j�B�܂�A����זE�ɂ�����n���i�ƃ~�g�R���h���A�ł̎_���I�����_���̗}����HIF-1�̊������i�ɂ���Ă����炳��Ă���B |
�y�W�N�����|�_�i�g���E���̓s���r���_�E���f�y�f�L�i�[�[��j�Q�����z
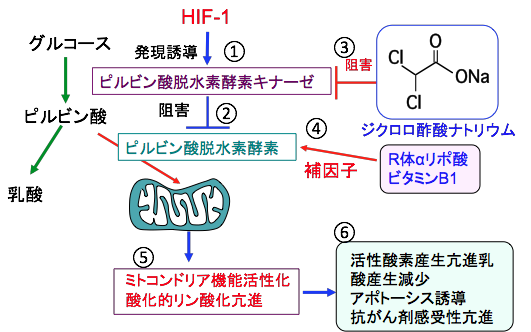
�W�N�����|�_�i�g���E���isodium dichloroacetate�j�͐|�_�iCH3COOH�j�̃��`����iCH3�j�̂Q�̐��f���q�����f���q�iCl�j�ɒu����������W�N�����|�_�iCHCl2COOH�j�̃i�g���E�����ł��B�\������CHCl2COONa�ɂȂ�܂��B �@
�W�N�����|�_�i�g���E���̓s���r���_�E���f�y�f�L�i�[�[��j�Q���邱�Ƃɂ���ăs���r���_�E���f�y�f�̊��������߂��p������܂��B�~�g�R���h���A�ُ̈�ɂ���Ӑ������A���_�A�V�h�[�V�X�A�S����]�̋����������̎��ÂȂǂɁA���i�Ƃ��ČÂ�����g�p����Ă��܂��B �@
�O�q�̂悤�ɂ���זE�ł�HIF-1�̊������i�ɂ���ăs���r���_�E���f�y�f�L�i�[�[�̊��������i���A�s���r���_�E���f�y�f�̊������ቺ���A�s���r���_����A�Z�`��CoA�ւ̕ϊ����j�~����Ă��邽�߁A�~�g�R���h���A�ł̃G�l���M�[�Y�����ቺ���Ă��܂��B �@
�����ŁA�W�N�����|�_�i�g���E���ł���זE�̃s���r���_�E���f�y�f�����������āA�s���r���_����A�Z�`��CoA�ւ̕ϊ��𑣐i����TCA��H���A���_�̎Y�����}�����܂��B����ɁA�_���I�����_���̉ߒ��Ŋ����_�f�̎Y���������A�_���X�g���X�̑���ɂ���Ă���זE�����łł���Ƃ�����p�@��������Ă��܂��i�}�W�j�B
|
�}�W�F��_�f�U�����q-1�iHIF-1�j�̓s���r���_�E���f�y�f�L�i�[�[�̔�����U�����āi�@�j�A�s���r���_�E���f�y�f�i�s���r���_���A�Z�`��CoA�ɕϊ�����j�̓�����j�Q����̂Łi�A�j�A�~�g�R���h���A�ł̎_���I�����_���ɂ��ATP�Y�����}������Ă���B�W�N�����|�_�i�g���E���̓s���r���_�E���f�y�f�L�i�[�[�̊�����j�Q���邱�Ƃɂ���ăs���r���_�E���f�y�f�̊��������߁i�B�j�AR��α���|�_�ƃr�^�~��B�P�̓s���r���_�E���f�y�f�̕���q�Ƃ��ē����i�C�j�A�s���r���_�E���f�y�f�̊��������߂ăs���r���_����A�Z�`��CoA�̕ϊ��𑣐i���ATCA��H�ł̑�ӂƎ_���I�����_���i����i�D�j�B�~�g�R���h���A�ł̎_���I�����_�������i����ƁA�����_�f�̎Y���������A���_�Y�����������A�A�|�g�[�V�X���N����₷���Ȃ��āA�R����܊������i����i�E�j�B |
����זE�ł͊����_�f�̔��������炵�Ď��ɂɂ������邽�߂ɁA�~�g�R���h���A�ł̎_���I�����_����}�����Ă��܂��B�W�N�����|�_�i�g���E���ł���זE�̃~�g�R���h���A�ł̑�ӂ𑣐i���Ċ����_�f�̎Y���𑝂₷�ƍR����܂Ŏ��ɂ₷���Ȃ�܂��B
�W�N�����|�_�i�g���E���P�Ƃł�����זE�����ł��邱�Ƃ��|�{�זE�⓮�������Ŗ��炩�ɂȂ��Ă��܂��B
�~�g�R���h���A�ł̊����_�f�̎Y���ʂ𑝂₵�����炢�ł���זE���{���Ɏ��ł���̂��Ƃ����^�₪�o�邩������܂��A�~�g�R���h���A�͍זE�̏d�ʂ�10%�ȏ���߂邭�炢��ʂɑ��݂���̂ŁA�~�g�R���h���A�ň�ĂɎ_���I�����_�������i����זE�����ł����邱�Ƃ��ł���ƍl�����Ă��܂��B
�|�{����זE�⓮���ڐA��ᇂ�p���������ŃW�N�����|�_�i�g���E���̍R��ᇍ�p�͏ؖ�����Ă��܂��B�Տ������ł��L����������Ă���A����̑�ֈ�Âł͗��p���銳�҂������Ă��܂��B �W�N�����|�_���R����ܑϐ������シ�邱�Ƃ���������Ă��܂��B
�l�Ԃ̏ꍇ�A�P���̏d1kg������10�`15mg�𐅂ɗn�����ĕ��p���܂��B�s���r���_�E���f�y�f������������ƁA���̍y�f�̕���q�ł����r�^�~��B1��R��α���|�_�����Ղ���̂��r�^�~��B1��R ��α���|�_�̕�[������p�\�h�ƌ��ʑ����ɕK�v�ł��B
|
�}�X�Fα���|�_�ɂ�R�̂�S�̂Ƃ����Q��ނ̌��w�ِ��́i�����ِ��́j�����݂���B�̓��Ő��������α���|�_��R�݂̂̂ŁAS�͓̂V�R�ɂ͑��݂��Ȃ��B�������Aα���|�_��l�H�I�ɍ��������R��50%�AS��50%�̃��Z�~�̂��ł���B�s���r���_�E���f�y�f�������������p��R�݂̂̂ŁA�t��S�̂�α���|�_�̓s���r���_�E���f�y�f�̊�����j�Q����B���������āA�W�N�����|�_�Ń~�g�R���h���A������������Ƃ��ɂ�R��α���|�_�łȂ���Ȃ�Ȃ��B |
�y���g�z���~����2-�f�I�L�V-D-�O���R�[�X�ƃW�N�����|�_�̑�������z
�Q-�f�I�L�V-D-�O���R�[�X�i2-DG�j�͉n��j�Q���邱�Ƃɂ����ATP�Y����j�Q���܂��B�o�����A�a������g�z���~���̓~�g�R���h���A�̌ċz�y�f��j�Q����ATP�̎Y����j�Q�����p������܂��B�ŋ߂̌����ł́A���g�z���~����2-DG�Ɠ��l�ɉn�y�f�̃w�L�\�L�i�[�[�̊�����j�Q�����p�����炩�ɂȂ��Ă��܂��B �@
���������āA2-DG�ƃ��g�z���~���p����ƁA����זE�̃G�l���M�[�Y����j�Q������ʂ����߂邱�Ƃ��ł��܂��B���ۂɁA�}�E�X�̈ڐA��ᇂ̎������f���ŁA2-DG�ƃ��g�z���~���p����Ƒ���I�ȍR��ᇌ��ʂ������邱�Ƃ��A�e�L�T�X��wMD�A���_�[�\������Z���^�[�������Ă��܂��B�iMol Cancer Ther. 10(12): 2350-2362, 2011�N�j
�|�{����זE��p���������ł́A2-DG�ʼnn��j�Q���Ă��A����זE�����ł����邾���̌��ʂ͓����܂��A���g�z���~�����ɓ��^����ƁA����זE�͎��ł��܂����B�l�X�Ȏ�ނ̂���זE���}�E�X�ɈڐA�������������ɂ����āA2-DG�ƃ��g�z���~���͂��ꂼ��P�Ƃł͍R��ᇌ��ʂ͎ア�̂ł����A���̂Q�p����Ƌ�����ᇏk�����ʂ��F�߂��Ă��܂��B �@
����זE�����B���邽�߂ɂ́A���B�̃V�O�i���ƁA�G�l���M�[�Y���ƕ��������̂��߂̍ޗ����K�v�ł��B���B�V�O�i���`�B�n�́A�C���X�����^�C���X�����l�������q-1�iIGF-1�j�Ƃ����̎�e�̂̌����ɂ���Ďh�������PI3K/Akt/mTORC1�`�B�n���d�v�ł��B
�@
���g�z���~���̓~�g�R���h���A�̌ċz���i�d�q�`�B�n�j�Ɖn�̃w�L�\�L�i�[�[��j�Q����ATP�Y����j�Q�����p������܂����A�����AMP�������v���e�C���L�i�[�[�iAMPK�j������������mTORC1�i�M���ރ��p�}�C�V���W�I�`����������-1�j�̊�����j�Q���邱�Ƃɂ���Ă���זE�̑��B��}�����܂��B
�@
����A2-DG�̓O���R�[�X�̉n�ƃy���g�[�X�E�����_�o�H�ł̑�ӂ�j�Q���邱�Ƃɂ���āA�G�l���M�[�Y���ƕ���������}�����A���̌��ʁA����זE�̑��B���}�����܂��B���Ȃ킿�A2-DG�ƃ��g�z���~���̓������^�́A����זE�̃G�l���M�[�Y���ƕ��������Ƒ��B�V�O�i���`�B�������I�ɑj�Q���邱�Ƃɂ���āA����זE�̑��B��j�Q���邱�Ƃ��ł���̂ł��B �@
���g�z���~���ɂ����_�A�V�h�[�V�X�������N��������p������܂��B���_�������āA���t���_���ɂȂ��Ԃł��B�傫�Ȃ���g�D������Ɠ��_�̎Y���������Ă��܂��B���_�A�V�h�[�V�X��h�����߂ɁA�̑��ł͓��_���O���R�[�X�i�u�h�E���j�ɕϊ��������V�������i���܂��B���g�z���~���͓��V����j�Q������ʂ�����̂ŁA���_�Y���̑���������ԂŃ��g�z���~���p����Ɠ��_�A�V�h�[�V�X���N�����₷���Ȃ�܂��B
���������āA����זE�̉n��}�����A�~�g�R���h���A�ł̎_�f�ċz�𑝂₷2-�f�I�L�V�O���R�[�X��W�N�����|�_�i�g���E����P�g���H�p����ƃ��g�z���~���ɂ����_�A�V�h�[�V�X�̔�����h�����Ƃ��ł��܂��B���ɃW�N�����|�_�i�g���E���͓��_�A�V�h�[�V�X�̎��ÂɌÂ�����g�p����Ă��܂��i�}10�j�B
|
�}10�F����זE�͉n�����i���ē��_�̎Y���������Ă���i�@�j�B���_�͊̑���t���₪��Ԏ��זE�œ��V���ɂ���ăO���R�[�X�ɕϊ�����Ă���i�A�j�B2-�f�I�L�V-D-�O���R�[�X�i2-DG�j�͂���זE�̉n��j�Q���i�B�j�A�W�N�����|�_�i�g���E���iDCA�j�̓s���r���_�E���f�y�f�����������ăs���r���_����A�Z�`��CoA�ւ̕ϊ��𑣐i����i�C�j�B���g�z���~���͓��V����j�Q����i�D�j�B���g�z���~���͌ċz����j�Q���āi�E�j�AATP�Y����ቺ�����i�F�j�A����Ƀ~�g�R���h���A�ł̊����_�f�̎Y���𑝂₵�Ď_���X�g���X�i����i�G�j�B�����̑g�ݍ����́A�_���X�g���X�����߁AATP�Y����ቺ�����āA����זE�̑��B��}�����A�זE����U������i�H�j�B2-DG��DCA�̓��g�z���~���ɂ����_�A�V�h�[�V�X��h���̂ŁA����p�����Ȃ��Ȃ�A�R��ᇊ��������߂邱�Ƃ��ł���B |
�~�g�R���h���A�̌ċz�y�f�����̂����g�z���~���őj�Q������ԂŃW�N�����|�_�ł���זE�̃~�g�R���h���A�̑�ӂi����A����זE�ɔ�r�I���ٓI�Ɏ_���X�g���X�����߂邱�Ƃ��ł��܂��B ���ۂɁA�W�N�����|�_�ƃ��g�z���~���̑�����ʂ�����������Ă��܂��B
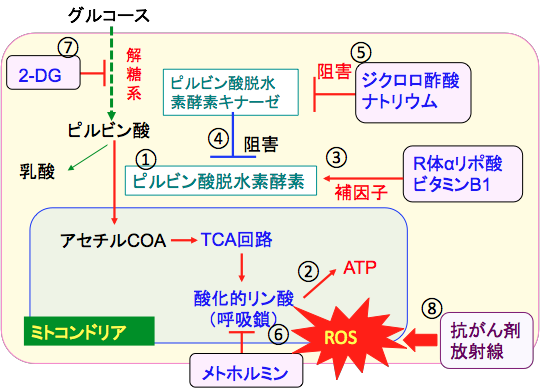
�ȏ�̂��Ƃ���A2-�f�I�L�V-D-�O���R�[�X�ƃ��g�z���~���ƃW�N�����|�_�i�g���E���ƁA�s���r���_�E���f�y�f�̕���q�̃r�^�~��B�P��R��α���|�_�̕��p�́A����זE�̉n��j�Q���A�~�g�R���h���A�����������邱�Ƃɂ���āA�R������Â���ː����Â̍R��ᇌ��ʂ����߂邱�Ƃ��ł��܂��i�}11�j�B
|
�}11�F�O���R�[�X���n�ő�ӂ���ăs���r���_�ɕϊ����ꂽ��A�s���r���_�E���f�y�f�ɂ���ă~�g�R���h���A���ŃA�Z�`��CoA�ɕϊ������i�@�j�B�A�Z�`��CoA�̓~�g�R���h���A����TCA��H�ƌċz�y�f�����̂ɂ�����_���I�����_���ɂ����ATP���Y�������i�A�j�BR��α���|�_�ƃr�^�~��B1�̓s���r���_�E���f�y�f�̊����ɕK�v�ȕ���q�ł���i�B�j�A�s���r���_�E���f�y�f�̓s���r���_�E���f�y�f�L�i�[�[�ɂ���ă����_������邱�Ƃɂ���Ċ������j�Q����Ă���i�C�j�B�W�N�����|�_�i�g���E���̓s���r���_�E���f�y�f�L�i�[�[��j�Q�����p������A���̌��ʃs���r���_�E���f�y�f������������i�D�j�B���g�z���~���͌ċz�y�f������I��j�Q���ă~�g�R���h���A�ł̊����_�f�̎Y���𑝂₷�i�E�j�B2-�f�I�L�V-D-�O���R�[�X�i2-DG�j�͉n��j�Q����ATP�Y���ƕ���������j�Q����i�F�j�B�R����܂���ː����Â��ŏI�I�ɂ���זE�����ł���Ƃ��Ɋ����_�f�ɂ���čזE�����U�������i�G�j�B���������āA�R������Â���ː����Â��s���Ƃ��ɁA2-DG�i2-�f�I�L�V-D-�O���R�[�X�j�A�W�N�����|�_�i�g���E���AR��α���|�_�A�r�^�~��B1�A���g�z���~���p����ƁA�R��ᇌ��ʂ��ł���B |
�y�זE���̍R�_���V�X�e���𗘗p���Ă���זE�͍R����ܒ�R���ɂȂ��z
�זE�ɂ́A�����_�f��L�ŕ����ɂ��Q����זE���g������i��d�g�݂�������Ă��܂��B
�Ⴆ�A�זE���Ŋ����_�f�̔����ʂ�������ƁA�זE�͊����_�f����������y�f�i�X�[�p�[�I�L�V�h�E�f�B�X���^�[�[�A�J�^���[�[�A�O���^�`�I���E�y���I�L�V�_�[�[�Ȃǁj�̔����⊈�������߂���A�t���[���W�J�������������O���^�`�I���Ȃǂ̍R�_�������̍��������߂��肵�āA�����_�f�̊Q�i�_���X�g���X�j���y�����悤�Ƃ��܂��B
�܂��A�O���^�`�I��S�g�����X�t�F���[�[�iGST�j�Ȃǂ��t�F�[�XII�i��Q���j ��ōy�f�ƌ������Ӎy�f�́A�l�X�Ȕ�������L�Q�����ʼn������p������܂��B
�זE�������_�f�┭������L�Q�Ȑ����i�R����܂���ː����܂ށj�ɂ���čU������ƁA�����̊����_�f�����y�f��R�_�������i�O���^�`�I����`�I���h�L�V���j��t�F�[�XII��ōy�f���זE���ɗU������i��`�q��������������Y���ʂ�������j�A�זE�����V�X�e���������܂��B
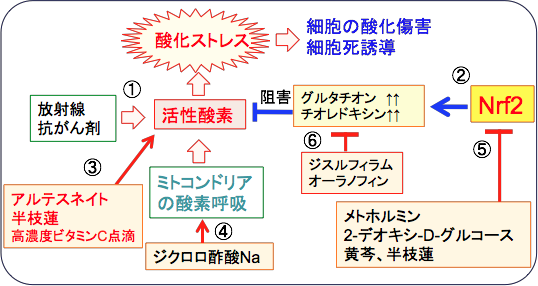
���̂悤�ȍזE���̖h��V�X�e���̊������ɒ��S�I�ȓ������s���Ă���̂�Nrf2�inuclear factor erythroid 2–related factor 2�j�Ƃ����]�ʈ��q�ł��B
|
�}12�F���ː���R����܂́A�����_�f�̎Y�������߁i�@�j�A�זE�̎_�����Q�������N�����āA�זE���B��}�����A�זE����U������i�A�j�B�_���X�g���X���y�����邽�߂ɓ]�ʈ��q��Nrf�Q�̊����i���i�A�j�A�X�[�p�[�I�L�V�h�E�f�B�X���^�[�[�iSOD�j��J�^���[�[��O���^�`�I���E�y���I�L�V�_�[�[�Ȃǂ̊����_�f�����y�f��O���^�`�I����`�I���h�L�V���Ȃǂ̍R�_�������̎Y�������߂āA�����_�f�ɂ��Q�i�_���X�g���X�j���y�����Ă���B���̍R�_���V�X�e���̘��i�ɂ���āA����זE�͕��ː���R����܂ɒ�R���ɂȂ�B |
�]�ʈ��q�Ƃ����͓̂���̈�`�q�̔����iDNA�̏���`�����ɕϊ����邱�Ɓj�߂��Ă���`�����ł��B �R�_���y�f��O���^�`�I���̎Y���Ɋւ���y�f��t�F�[�X�Q��ōy�f�̈�`�q�̔������ߗ̈�ɂ́A�R�_�������G�������g�iantioxidant response element�FARE�j�Ƃ����̈悪�����āA���̕�����Nrf2����������Ƃ����̈�`�q�̓]�ʂ����i�����̂ł��B
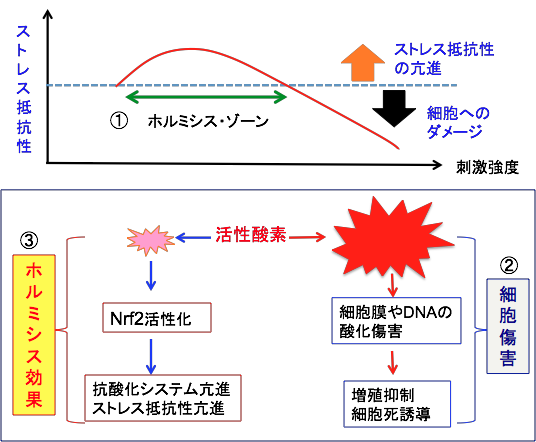
�z���~�V�X�ihormisis�j�����Ƃ������̂�����܂��B�̂ɗL�Q�Ǝv���Ă�����ː��⊈���_�f�₠���̔��������A���ʂł���Α̂��h����������œ����āA�����̊Q�ɑ����R�͂����܂�Ƃ������ʂł��B�h����X�g���X������Ԃ����ƁA���̂͂����ɑ��ēK������悤�ɑ̂��������邽�߁A���̎h����X�g���X�ɑ��Ē�R���ɂȂ�ƍl������̂ł����A�R�_���͂��ŗ͂̑����̃��J�j�Y���Ɋւ��Ă͓]�ʈ��q��Nrf2�̊��������֗^���Ă��܂��B
�����܁i�_��j�̃p���R�[�g�͊����_�f�������܂��B������l�X�ȔZ�x�̃p���R�[�g�̓������|�n�ň�ĂāA���̎�����������������������܂��B
�p���R�[�g�̔Z�x���ɂ߂ĒႢ�i0.005mM�ȉ��j�Ǝ����ɉe���͋y�ڂ��܂��A�Z�x��0.01mM����0.5mM�̏ꍇ�́A�������ő��60%���炢�������܂��B1mM�ȏゾ�Ƌt�Ɏ����͒Z�k���܂��B
�y�x�̎_���X�g���X�͎����������A���x�̎_���X�g���X�̓_���[�W��^����̂Ŏ����͒Z�k�����Ƃ������ʂł��B
���w�������̌����ł��z���~�V�X���ʂ��F�߂��Ă��܂��B�̓��Ŋ����_�f�������Ĕ������p�������悤�ȕ��������ʂ������^����ƁA�������Ĕ����}�����邱�Ƃ�����܂��B����́A�y�x�̎_���X�g���X�ɑ��āA�K�������Ƃ��đ̂̒��̍R�_���y�f���������邽�߂ł���ƍl�����Ă��܂��B���̏ꍇ���ANrf2�̊��������d�v�Ȗ�����S���Ă��܂�.
����Nrf2�͗l�X�ȃX�g���X��L�Q��������זE������Ă���̂ŁA�זE�ɂƂ��Ă͏d�v�Ȗh��V�X�e���ł��B
|
�}13�F�����ɑ��Ēʏ�L�Q�ȍ�p�������h�����A���ʂł���t�ɗL�v�ȍ�p�i�X�g���X��R���̘��i�j�ɂȂ�Ƃ������ۂ��u�z���~�V�X�iHormesis�j�v�ƌ����i�@�j�B�����_�f�͑�ʂɔ�������ƍזE����DNA�̎_�����Q���瑝�B�}����זE���U���Ȃǂ̍זE���Q��p�����߂��i�A�j�B�������A�K�x�Ȋ����_�f�̎Y���́A�]�ʈ��q��Nrf2�̊����������z���~�V�X���ʂɂ���āA�R�_���V�X�e���̘��i��X�g���X��R���̘��i��U������i�B�j�B |
����זE�ł�Nrf2�̊��������ɍ����Ȃ��Ă���A������Nrf2�̊�������������זE�قǎ��Âɒ�R���ŗ\�オ�����Ƃ������ʂ������Ă��܂��B
�܂�A����זE�́A�R����܂���ː����Â��玩������邽�߂�Nrf2�𗘗p���Ă���Ƃ������Ƃł��B
����זE�ɂ�����Nrf2�̓�����j�Q���邱�Ƃ��ł���A�R����܂���ː����Â̌����ڂ����߂邱�Ƃ��ł��܂��B�|�{�זE�⓮�������̒i�K�ł����A�R������Â���ː����Â�Nrf2�j�Q�܂p����ƍR��ᇌ��ʂ����܂邱�Ƃ�����Ă��܂��B
�yNrf2�͍זE�̍R�_���͂����߂��z
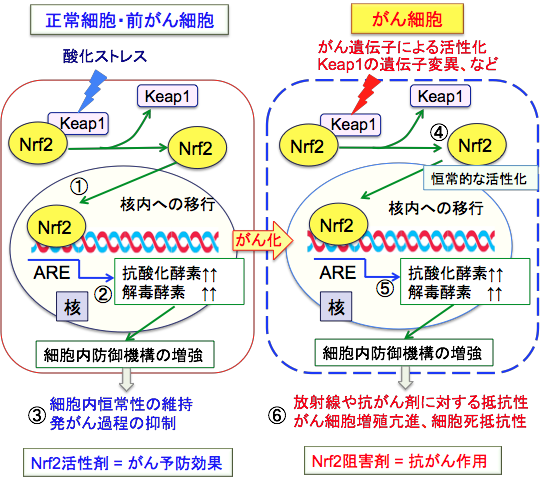
�]�ʈ��q��Nrf2�inuclear factor erythroid 2–related factor 2�j�́A�R�_���@�\���ŋ@�\�����l�X�Ȉ�`�q�̓]�ʂ�U�����܂��B
�����_�f���H���Ɋ܂܂��e�d�q�������ɂ������炳���_���X�g���X�́CDNA��^���p�N���⎉���Ȃǂ̐��̍����q���_�����邱�Ƃŏ��Q��^���A����ⓜ�A�a��t��������_�o�ϐ������ȂǗl�X�Ȏ����������N���������ɂȂ�܂��B
���̂悤�Ȏ_���X�g���X�ɑ���h��@�\�ɂ����ďd�v�ȋ@�\��S���Ă���̂��AKeap1-Nrf2�V�X�e���ł��D ����זE�ɂ����Ď_���X�g���X�̖����ł́ANrf2��Keap1�iKelch-like ECH-associated protein 1�j�Ƃ����^���p�N���Ɍ������邱�Ƃɂ�胆�r�L�`�������A�������Ă��܂��B
�������A�_���X�g���X�ɂ��炳����Keap1�̃V�X�e�C���c��C�����č\�����ω����AKeap1����Nrf2������Ċj�ֈڍs���A��`�q�㗬�ɑ��݂����R�_���܉����z��ARE�iantioxidant response element�j�Ɍ������邱�Ƃɂ���āA����ARE�z������l�X�Ȉ�`�q�i�R�_���y�f��O���^�`�I���̍����Ɋ֗^����y�f��t�F�[�XII��ōy�f�Ȃǁj�̔�����U�����A�R�_�����łɊւ���^���p�N������q�̍��������߂܂��B
���̂悤�ȍ�p�ɂ���āA����זE��O����זE�ɂ����ẮANrf2�̊����i����Nrf2�����܂͔�����ߒ���}������̂ŁA����\�h�����Ƃ��ĔF������Ă��܂��B���̂悤��Nrf2�̊��������������\�h�����Ƃ����X���t�H���t�@�����N���N�~�������X�x���g���[���Ȃǂ�����Ă��܂��B
����A����זE�ɂ����ẮANrf2���P��I�Ɋ��������Ă���ANrf2������j�Q����Ƃ���זE�͎��ɂ₷���Ȃ邱�Ƃ�����Ă��܂��B �q�g�̔x����ȂǑ����̌Ō`��ᇂ�Nrf2�@�\�ُ̈혴�i���������Ă��܂��B�����āANrf2�̊����������Ǝ��Âɒ�R���ŗ\�オ�s�ǂł��邱�Ƃ�����Ă��܂��B
Nrf2�͉�ōy�f�A�R�_���^���p�N���A��ܔr�o�g�����X�|�[�^�[�Ȃǂ̈�`�q���I�Ɋ��������邱�Ƃɂ��A����זE�̍R����ܑϐ��ƕ��ː��ϐ��������炵�܂��B
����ɁANfr2�̓O���R�[�X��O���^�~���̑�ӂ�ω������čזE���B�ɗL���ȓ��������𑣐i���܂��B
Nrf2�͂���זE�̃y���g�[�X�����_�o�H�����������āA�j�_��NADPH�̎Y���𑝂₷���ƂōזE���B�i���܂��BNADPH�̓O���^�`�I���̍����ɂ��K�v�ł��B
�����̂���זE�ł́A�_���X�g���X�̗L���Ƃ͊W�Ȃ�Nrf2�̔����ʂƊ��������i���Ă��܂��B���̗��R�́ARAS��MYC�Ȃǂ̂����`�q��Nrf2�����������Ă��邩��ł��B Nrf2�̊�����}������Keap1�̈�`�q�ψقɂ����Nrf2������������ꍇ���������Ă��܂��B
�P��I�Ɉ��艻����Nrf2�͎_���X�g���X��R����܁^���ː����Âɑ����R�������A����ɐϋɓI�ɍזE���B�𑣐i���邱�ƂɂȂ�܂��B
�R������Â���ː����Âɂ���Ċ����_�f��������ƁA����זE�͂����Nrf2�̗ʂ𑝂₵�Ē�R�����l�����Ă��܂��B
���̂悤��Nrf2�̊������͂���זE�����Âɑ����R�����l�����郁�J�j�Y���Ƃ��ďd�v�ł��B���������āA����זE�̂�����Nrf2�̋@�\�j�Q�́A�R������Â̗L���Ȑ헪�ƂȂ�܂��i�}14�j�B
|
�}14�FKeap1-Nrf2�V�X�e���͎_���X�g���X��L�Q�����ɑ���h��@�\�ɂ����ďd�v�Ȗ�����S���Ă���B�]�ʈ��q��Nrf2�͍זE����Keap1�ɂ���ĕ��������i����邱�Ƃɂ���Ċ������}������Ă���B����זE�ł͎_���X�g���X�������ƁAKeap�P�̍\�����ω�����Nrf2���痣��A�t���[�ɂȂ���Nrf2���j���Ɉڍs���āi�@�j�A�R�_���y�f���ōy�f�̈�`�q�̏㗬�ɑ��݂���R�_���܉����z��ARE�iantioxidant response element�j�Ɍ������āA�����̈�`�q�̔����i����i�A�j�B���̌��ʁA����זE�ł͊����_�f�┭��������ł��邱�ƂɂȂ�̂ŁANrf2�̊����𑣐i������́i��Nrf2�����܁j�͂���\�h���ʂ�����i�B�j�B ����A����זE�ɂ����ẮAKRas��cMyc�Ȃǂ̂����`�q�̊�������Keap1�̈�`�q�ψقȂǂɂ����Nrf2�͍P��I�Ɋ��������Ă���i�C�j�A�R�_���y�f���ōy�f�̎Y���⊈�������i���Ă���B���̂��߁A����זE�͎_���X�g���X��זE���Q�������ɑ����R�������i���Ă���i�D�j�A���ꂪ�R����܂���U�����Âɑ����R���̌����ɂȂ��Ă���i�E�j�B���������āA����זE�ɂ����ẮANrf2�̊�����j�Q������́i��Nrf2�j�Q�܁j�����Âɖ𗧂B���̂悤�ɁA����זE�i�O����זE���܂ށj�Ƃ���זE�Ƃł�Nrf2�ɑ���Ή��͑S���قȂ�B |
���A�a���t�ǂ▝���ǐ��x�����ȂǑ����̎_���X�g���X�Ɗ֘A���鎾���̗\�h�⎡�Â̂��߂�Nrf2�̊������܂��Տ����p�Ɍ����ĊJ������Ă��܂��B
����A����̎��ÂɂȂ��Nrf2�̑j�Q�܂��L�v�ɂȂ�\���������ƍl�����Ă��܂��B ����זE��Nrf2��j�Q����ƁA
�@����זE���ł̊����_�f��̒~�ςɂ���ăA�|�g�[�V�X�i�זE���j���U���ł���
�A�R������Â���ː����Â̌����ڂ����߂邱�Ƃ��ł���
�B����זE�ł̕��������i���������j��j�Q���邱�Ƃɂ���đ��B��j�Q�ł���A
�Ȃǂ̌��ʂ����҂ł��܂��B
�������A�S�g���^�̏ꍇ�́A����g�D�ȊO�̐���g�D�ł̎_���X�g���X���������コ���錇�_������܂��B�܂�A����זE���ٓI��Nrf2��j�Q�ł�����@������A���Âɖ𗧂��܂��B
�y2-�f�I�L�V-D-�O���R�[�X�ƃ��g�z���~���͂���זE���ٓI��Nrf2������j�Q�����z
Nrf2�̓]�ʊ���������R�_���y�f���ōy�f�̔����ɂ̓O���R�[�X�̋������K�v�ł���Ƃ���������܂��B
Glucose availability is a decisive factor for Nrf2-mediated gene expression.�i�O���R�[�X�̋�����Nrf2�������`�q�����̂��߂̌���I�ȗv���ł���jRedox Biol. 2013 Jun 21;1(1):359-65.
���̘_���ł́A�X���t�H���t�@����Nrf2��������������@��ANrf2��j�Q����Keap1��`�q���@�\������������@�i��`�q�m�b�N�A�E�g�j��Nrf2�̊����i������@�Ŏ������Ă��܂��B
Nrf2������������ƃO���R�[�X�̎捞�݂������A�y���g�[�X�����_�o�H�ł�NADPH�̎Y���������邱�Ƃ�������Ă��܂��B
�����āA�O���R�[�X�̋�����捞�݂�j�Q���邩�A���邢�̓y���g�[�X�����_�o�H��j�Q����NADPH�̎Y����j�Q����ƁANrf2�������`�q�������}������A�R�_���y�f���ōy�f�̔������}������邱�Ƃ�������Ă��܂��B
�܂�A����������P�g���H��Q−�f�I�L�V�O���R�[�X�ȂǂŁA����זE�ɂ�����O���R�[�X�̎捞�݂�y���g�[�X�����_�o�H��}��������@�́A�O���R�[�X�̎捞�݂����i���Ă��邪��זE�ɑI��I��Nrf2�̊�����j�Q�ł���\��������܂��B
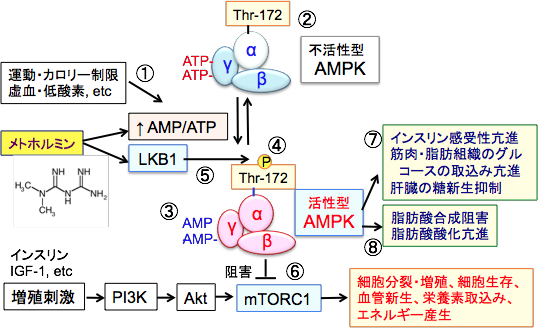
���g�z���~���imetformin�j�́A���E���łP���l�ȏ�̂Q�^���A�a���҂Ɏg���Ă���r�O�A�i�C�h�n�o�������~���܂ł��B �O�q�̂悤�ɁA���g�z���~���̓~�g�R���h���A�̌ċz���̍ŏ��̃X�e�b�v�ł����ċz�y�f������I��j�Q���邱�Ƃ����炩�ɂȂ��Ă��܂��B���̌��ʁA�~�g�R���h���A�ł�ATP�Y�����������AAMP:ATP�̔䂪�㏸���AAMP�������v���e�C���L�i�[�[�iAMPK�j������������܂��B
����������AMPK�́A�̑��̓��V����}�����A�i���A���i�ł̃O���R�[�X���p�𑣐i���Č�����ቺ�����܂��B ���Ȃ킿�A���g�z���~���̌����~����p�̓~�g�R���h���A�ɂ�����ATP�Y���̑j�Q�ɂ���đ̓���ATP�ʂ��������邽�߂ł��B�̂�ATP�𑝂₷���߂ɁA�O���R�[�X�̕����i�ى��j�𑣐i���A���V���i�����j��}������̂ŁA�������ቺ���܂��B ����ɁAAMPK�̊������͍זE���B�𑣐i����mTORC1�i�M���ރ��p�}�C�V���W�I�`����������-1�j�̊�����j�Q���邱�Ƃɂ���Ă���זE�̑��B��}�����܂��B�i�}15�j
|
�}15�FAMP�������v���e�C���L�i�[�[�iAMPK�j�͍זE�̃G�l���M�[��ӂ߂�����q�Ƃ��ďd�v�Ȗ�����S���Ă���BAMPK�͒�O���R�[�X���_�f�⋕���ȂǍזE��ATP�������͊�������悤�ȃX�g���X�ɉ������Ċ����������i�@�j�BAMPK�͐G�}��p������α�T�u���j�b�g�ƁA���ߍ�p������β�T�u���b�g��γ�T�u���j�b�g����\�������w�e���O�ʑ̂Ƃ��đ��݂���i�A�j�Bγ�T�u���j�b�g�ɂ�ATP���������Ă��邪�AATP���͊�����AMP/ATP�䂪�㏸����ƁAγ�T�u���j�b�g�Ɍ������Ă���ATP��AMP�ɒu�������i�B�j�B���̌��ʁA�A���X�e���b�N���ʁi�y�f�̗��̍\�����ω����邱�Ɓj�ɂ���Ă��̕����̂͒����x�i2�`10�{���x�j�Ɋ���������A�㗬�Ɉʒu�����v��AMPK�L�i�[�[�ł���LKB1�ɑ��Đe�a���������Ȃ�ALKB1�ɂ����α�T�u���j�b�g�̃X���I�j��-172�iThr-172�j�������_�������ƁA�y�f�����͍ő�Ɋ����������i�C�j�B LKB1�̓Z�����E�X���I�j���L�i�[�[�ŁAAMP�������v���e�C���L�i�[�[�iAMPK�j�������_�����Ċ���������i�D�j�B�����_�����ꂽAMPK��mTORC1�i�M���ރ��p�}�C�V���W�I�^���p�N�������̂P�j��}�����A�^���p�N���⎉�b�_�̍�����}�����āA����זE�̑��B��}������i�E�j�B������AMPK�̓C���X���������i�A�ؓ��E���b�g�D�̃O���R�[�X�̎捞�ݘ��i�A�̑��̓��V���}���ɂ���Č�����ቺ������i�F�j�B����ɁA���b�_������j�Q���A���b�_�̎_���i����i�G�j�B |
���g�z���~��������זE��Nrf2������}�����čR��ᇍ�p���������Ƃ����炩�ɂȂ��Ă��܂��B �ȉ��̂悤�ȕ�����܂��B
Metformin inhibits heme oxygenase-1 expression in cancer cells through inactivation of Raf-ERK-Nrf2 signaling and AMPK-independent pathways.�i���g�z���~����Raf-ERK-Nrf2 �V�O�i���`�B�n��AMPK��ˑ��I�o�H�̑j�Q�ɂ���Ă���זE�̃w���I�L�V�Q�i�[�[-1��j�Q����jToxicol Appl Pharmacol. 2013 Sep 1;271(2):229-38.
�w���I�L�V�Q�i�[�[-1�́A�w���iheme�j���r���x���W���ibiriverdin�j�ƈ�_���Y�f�iCO�j�ƗV���S�iFe�j�ɕ�������y�f�ł��B�r���x���W���i�ΐF�̐F�f�j�́A�r���x���W���E���_�N�^�[�[�ibiriverdin reductase�j�ɂ��A�r�����r���ibilirubin�F���t�̎��̉��F�̐F�f�j�ɕ�������܂��B���̃w�������ɂ��Y�����r���x���W���ƃr�����r���ɂ͋��͂ȍR�_����p������A�_���X�g���X�ɂ��זE���Q��}�����܂��B
���g�z���~����Raf-ERK�V�O�i���`�B�n��}����Nrf2�̔����ʂ����炷���ʂ����邱�Ƃ���Ă��܂��B
Metformin Sensitizes Non-small Cell Lung Cancer Cells to an Epigallocatechin-3-Gallate (EGCG) Treatment by Suppressing the Nrf2/HO-1 Signaling Pathway.�i���g�z���~���́ANrf2 / HO-1�V�O�i���`�B�o�H��}�����邱�Ƃɂ��A�זE�x����זE���G�s�K���J�e�L��-3-�K���[�g���ÂɊ��ɂ���B�jInt J Biol Sci. 2017 Nov 27;13(12):1560-1569.
�Β����̎�v�|���t�F�m�[���ł����G�s�K���J�e�L��-3-�K���[�g�iEGCG�j�́A�R�����p��L���܂��B NF-E2�֘A���q2�iNrf2�j/�w���I�L�V�Q�i�[�[-1�iHO-1�j�V�O�i���`�B�o�H�̊������́AEGCG�ɑ���זE��R���������N�����܂��B���̌����ł́A���g�z���~����HO-1������j�Q���AEGCG�̍R��ᇌ��ʂ����邱�Ƃ𖾂炩�ɂ��Ă��܂��B
���g�z���~���́AEGCG�i100μM�j�ɂ���ėU������銈���_�f��̎Y���𑝂₵�A�A�|�g�[�V�X��U�����܂����B ����ɈڐA��ᇂ�p�������������ŁA���g�z���~����EGCG�̕��p���^�͎�ᇂ̏k�����ʂ𑊏�I�ɑ������܂����B
���J�j�Y���I�ɂ́AEGCG�Ŋ��������ꂽNrf2 / HO-1�o�H���A���g�z���~����Nrf2��SIRT1�ˑ����E�A�Z�`�����ɂ���đj�Q���܂����B����ɁA���g�z���~���́A�����I��NF-kB�o�H�����SIRT1�����i���܂����B
����Ɣ�r���āAEGCG�ƃ��g�z���~���Ƃ̑g�ݍ��킹�͐���Ȕx���זE�ɑ��ĂقƂ�lje����^���܂���ł����B
�ȏ�̌��ʂ���A���g�z���~���́ANrf2 / HO-1�V�O�i���`�B�o�H��}�����邱�Ƃɂ��A�זE���x����זE��EGCG�̍R�����p�ɑ��銴�����߂��Ƃ�������Ă��܂��B
�G�s�K���J�e�L��-3-�K���[�g�iEGCG�j�͗Β����̎�v�ȃ|���t�F�m�[���ł��BEGCG�͂���זE�ɍ�p���Ď_���X�g���X�������N�����āA�R�����p�����܂��B
�|���t�F�m�[���͂���זE�ɑ��Ď_���܂Ƃ��č�p���đ��B�}����זE���U���̍�p������Ƃ����l�����ŋ߂͎嗬�ɂȂ����܂��BEGCG�̍R��ᇍ�p�ɑ��āA����זE��Nrf2/�w���I�L�V�Q�i�[�[-1�iHO-1�j�V�O�i���`�B�o�H�̘��i�ɂ���Ē�R���悤�Ƃ��܂��B ����ɑR���āA���g�z���~����Nrf2/HO-1�V�O�i���`�B�o�H��}������HO-1������j�Q���AEGCG�̍R��ᇌ��ʂ����邱�Ƃ������Ă��܂��B
���g�z���~���͂���זE�ɑ���Nrf2/HO-1�o�H��}�����邱�Ƃɂ���āA�R�_���V�X�e������̉������p�����҂ł���悤�ł��B
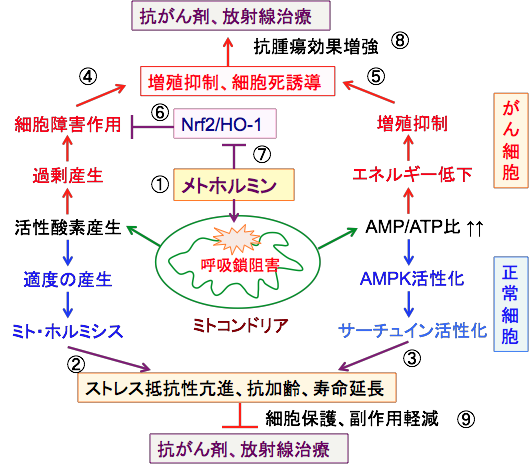
����זE�Ƃ���זE�̃��g�z���~���̍�p�̈Ⴂ�́A�~�g�R���h���A�ɂ����銈���_�f�̗ʂɂ��\��������܂��B����זE�͍זE�ɔ�ׂĊ����_�f���������₷���ɂ���A�����_�f�̓K�x�Ȕ����̓z���~�V�X���ʂɂ���ėL�v�ȍ�p�������A����זE�ł͑�ʂ̊����_�f���o�₷���̂ŁA�זE���Q���D�ʂɂȂ�\��������܂��i�}16�j�B
|
�}16�F���g�z���~���̓~�g�R���h���A�̌ċz����j�Q���Ċ����_�f�̎Y�������߁AATP�Y����ቺ������i�@�j�B���̍�p�͐���זE�ɑ��ẮA�K�x�Ȋ����_�f�̎Y�����~�g�z���~�V�X�̍�p�ɂ���ăX�g���X��R�������߁i�A�j�AAMP/ATP��̏㏸��AMPK�iAMP�������v���e�C���L�i�[�[�j��T�[�`���C�������������āA�R����i����֘A�����̗}���j������������ʂ�����i�B�j�B����A����זE�ɂ����Ă̓~�g�R���h���A�Ŋ����_�f���Y������₷���ɂ���B���̌��ʁA�ċz���̑j�Q�͊����_�f�̎Y�����ߏ�ɂȂ��Ď_�����Q�������N�����i�C�j�A�G�l���M�[�ቺ�ɂ���đ��B��}������i�D�j�B�_���X�g���X�̘��i�ɑ��Ă���זE��Nrf2/HO-1�n�����������Ď_�����Q���y�����悤�ƒ�R����i�E�j�B���g�z���~����Nrf2/HO-1�n��j�Q���邱�Ƃɂ���Ă���זE�̍R�_���V�X�e������̉�����i�F�j�B�_�����Q�ɂ���Č��ʂ�����R����܂���ː����Âɑ��āA���g�z���~���͂���זE�ɑ��Ă͍R��ᇌ��ʂ����߁i�G�j�A����זE�ɑ��Ă͕ی�I�ɍ�p���ĕ���p���y������i�H�j�B |
���g�z���~�����R����܂���ː����Â̌����ڂ����߂��Ƃ͑����̕�����܂��B����ɁA�l�X�ȃ��J�j�Y���ōR����܂���ː����Â̕���p�̌y���ɂ��L���ł��邱�Ƃ�����Ă��܂��B
����ɁA2-�f�I�V�L-D-�O���R�[�X��W�N�����|�_�ƕ��p����ƁA�R��ᇌ��ʂ����܂邱�Ƃ������������Ă��܂��B����̕W�����Â̌����ڂ����߂�ړI�����g�z���~���{�Q−�f�I�L�V-D-�O���R�[�X�{�W�N�����|�_�i�g���E���i+�r�^�~��B1�AR��α���|�_�j�̑g�����́A�����Ɨ��p����Ă��ǂ��Ǝv���܂��B
�y�����_�f�̎Y���𑝂₷�����z
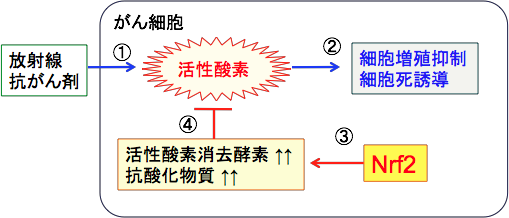
����זE�̎_���X�g���X�����߂Ă���זE�����ł����鎡�Ö@�́uoxidation therapy�i�_�����Áj�v�ƌĂ�Ă��܂��B
����זE�ł̓~�g�R���h���A�̋@�\�ُ�Ȃǂɂ���Ď_�f�ċz���s���Ɗ����_�f�̎Y�������܂�܂��B ����זE�͎_���X�g���X�����߂����Ȃ��̂ŁA�~�g�R���h���A�ł̑�ӂ�}�����A�_�f���g��Ȃ��n�ł̑�ӂi�����Ă��܂��B
���������āA�n��}�����~�g�R���h���A�ł̎_���I�����_���i�_�f���g�����G�l���M�[�Y���j�i����A����זE���̎_���X�g���X��\���I�ɍ��߁A����זE�����ł����邱�Ƃ��ł��܂��B
���ː����Âƈꕔ�̍R����܁i�r���u���X�`���A�V�X�v���`���A�}�C�g�}�C�V��C�A�h�L�\���r�V���A�J���v�g�e�V���Ȃǁj�́A����זE�Ɏ_�����Q�������N�����čזE�Ƀ_���[�W��^���Ď��ł����܂��B���̑��̍R����܂��A�זE���������N�����Ƃ��Ɋ����_�f���g���܂��B
���������āA���ː����Â�R������Â̍Œ��́A�R�_����p�̂���T�v�������g�̐ێ�͍זE����j�Q���܂��B����A�_���X�g���X������A���ː����Â�R������Â̌����ڂ����߂邱�Ƃ��ł��܂��B
�������N�̓����Ƃ��āA����זE�̎_���X�g���X�����߂鎡�Â����ڂ���Ă��܂��B
��{�́A�n��j�Q�i2-DG�j���A�~�g�R���h���A�̊������Ȃǂɂ���Ċ����_�f�̎Y�������߁i�W�N�����|�_�A���g�z���~���j�A����זE�̍R�_���V�X�e����j�Q������@�i���g�z���~���A������A�W�X���t�B�����A�I�[���m�t�B���j�̕��p���L���ł��B ����ɂ���זE�Ɏ_���X�g���X�����߂��P�g���H�A�A���e�X�l�C�g�A���Z�x�r�^�~��C�_�H�A���}�@�̐�����p����ƁA����זE�Ɏ_�����Q��^���āA���B��}�����A�זE����U���ł��܂��B
���g�z���~���⍂�Z�x�r�^�~��C�_�H��A���e�X�l�C�g�⊿�����W�N�����|�_�Ȃǂ��R������Â���ː����Âƕ��p���čR��ᇌ��ʂ����߂鍪���́u�����_�f�̎Y�������߂邩���v�ƍl����̂������ɍ����Ă���Ǝv���܂��i�}17�j�B
|
�}17�F���ː���R����܂́A�����_�f�̎Y�������߂Ď_���X�g���X�i���A�זE�̎_�����Q�������N�����čזE����U������i�@�j�B�_���X�g���X���y�����邽�߂ɓ]�ʈ��q��Nrf�Q�̊����i���A�O���^�`�I����`�I���h�L�V���V�X�e���i���Ċ����_�f�ɂ��_�����Q�ɒ�R����i�A�j�B�A���e�X�l�C�g�A���}�@�i�͂��j�A���Z�x�r�^�~��C�_�H�͍זE���̊����_�f�̎Y�������߂�i�B�j�B�W�N�����|�_�i�g���E���̓~�g�R���h���A�̎_�f�ċz�i���Ċ����_�f�̎Y�������߂�i�C�j�B���g�z���~����2-�f�I�L�V-D-�O���R�[�X�≩?�┼�}�@��Nrf2�̊�������j�Q����i�D�j�B�W�X���t�B������I�[���m�t�B���̓O���^�`�I����`�I���h�L�V���ɂ��R�_���V�X�e����j�Q���邱�Ƃɂ���Ď_���X�g���X�����߂�i�E�j�B������g�ݍ�����ƁA����זE�Ɍ����I�Ɏ_���X�g���X�����āA����זE�̑��B��}���ł��A�R����܂���ː����Â̍R��ᇌ��ʂ��ł���B |
�y���זE�̃}�[�J�[�̃A���f�q�h�E���f�y�f�̓V�X�v���`���ϐ��Ɋ֗^�����z
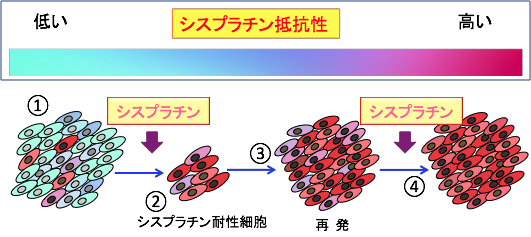
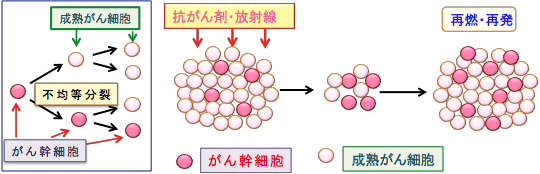
���זE�icancer stem cell�j����ᇎn���זE�itumor initiating cell�j�Ƃ��Ă�A����זE�ݏo�����ƂɂȂ�זE�ł���A����g�D���ɏ����i�������x�j���݂��Ă��܂��B �ʏ�̍R������Â���ː����Âɑ��āA���n��������זE�͎��ł��₷���̂ł����A���זE�͗l�X�ȋ@���Œ�R���������܂��i�}18�j�B
|
�}18�F����g�D�ɂ͂��זE (cancer stem cells)�ƌĂ��זE�����݂��āA�ʏ�̂���זE�i���n����זE�j���������Ȃ��炪��g�D���\�����Ă���B���זE�͎��ȕ������s������A�s�ϓ�����ɂ��ꕔ�����ȕ����̃T�C�N�������E���ĕ������ʏ�̂���זE�ƂȂ��Ă���B���n����זE�͍R����܂���ː��Ŏ��ł��₷�����A���זE�͎��ł��ɂ����̂ōR������Â���ː����ÂŐ����c��B���זE�͎�ᇌ`���\�����̂ŁA�����c�������זE�����B���čĔ���ĔR���N����B���Â��J��Ԃ��Ǝ��Ò�R���̂��זE�������A���Ò�R�������i����B���זE���A�|�g�[�V�X��R���ɂȂ��Ă��郁�J�j�Y����j�Q����R����܂���ː����Â̌��ʂ����߂邱�Ƃ��ł���B |
���זE�������c��A����͂�����ĔR�E�Ĕ����܂��̂ŁA���זE�̕��ː�����R����܊������߂���@�̊J�����d�v�ƍl�����Ă��܂��B
���̕��@�̈�Ƃ����A���f�q�h�E���f�y�f�j�Q�������ڂ���Ă��܂��B �A���f�q�h�E���f�y�f�͂��זE�ɑ����������A���̍y�f�̔�������������قǑ��B�������\�オ�������Ƃ�����Ă��܂��B
�A���f�q�h�E���f�y�f�͂��זE�ɑ����������A���זE�̃}�[�J�[�Ƃ��Ďg�p����Ă��܂��B���̃A���f�q�h�E���f�y�f���R����܂̃V�X�v���`���ϐ��ɏd�v�ȓ��������Ă��邱�Ƃ������̌����Ŏ�����Ă��܂��B �ȉ��̂悤�Ș_��������܂��B
Targeting the cancer stem cell marker, aldehyde dehydrogenase 1, to circumvent cisplatin resistance in NSCLC�i�זE���x����ɂ�����V�X�v���`���ϐ����������邽�߂ɁA���זE�}�[�J�[�ł���A���f�q�h�E���f�y�f1��W�I�Ƃ���jOncotarget. 2017 Sep 22; 8(42): 72544–72563.
�y�v�|�z
�זE�x����͂��̑������߁A���Ñt�������Ⴍ�A�\��͈�ʓI�ɕs�ǂł���B���q�W�I��Ŏ��Â��\�ȓ��ٓI�ȓˑR�ψق����݂��Ȃ��ꍇ�A�V�X�v���`�����x�[�X�ɂ����R������Â��A���̎����̎��Âɂ����ďd�v�Ȗ������ʂ����B
�������Ȃ���A����זE���V�X�v���`���ɑϐ����l�����邱�Ƃ��A���̍זE���Q���̎g�p�ɂ����Ď�v�Ȏ��Ï�̉ۑ�ƂȂ��Ă���B ���̖�ܑϐ��̃��J�j�Y�����𖾂��邱�Ƃ́A�זE���x���҂ɂ�����V�X�v���`���ɑ��銴�����߂�V�K��܂̊J���������炷�\��������B
���̌����ł́A�זE���x����̍R����ܒ�R��������������@�Ƃ��āA���זE�ɂ������A���f�q�h�E���f�y�f1�iALDH1�j�̊������^�[�Q�b�g���邱�Ƃ����������B
�זE���x���җR���̎�ᇂ́A�����̑��\�����זE�֘A��`�q�̔���������F�߂��B�V�X�v���`���̓��^�́A�V�X�v���`��������ёϐ��̔זE���x����זE���ɂ����āAALDH1�z���̂���זE�W�c�̏o���܂��͑����U�����A�V�X�v���`���ϐ�������ɍ��߂��B
Aldefluor�A�b�Z�C�����FACS���͂�p����ALDH1�z���זE��P�����A���זE�����Ɋւ��ĉ�͂����B ALDH1�z���זE�݂̂��A�s�ϓ�����A�V�X�v���`���ϐ������in vitro�ł̊��זE���q�̔����̑������������B
NOD / SCID�}�E�X�ɂ�����َ�ڐA�����ł́A������ALDH1�z������זE�W�c�����ALDH1�A������זE�W�c��������I�Ȏ�ᇌ`�����������B �W�G�`���A�~�m�x���Y�A���f�q�h���邢���W�X���t�B������ALDH1��j�Q����ƁA�V�X�v���`����R���̔x����זE���V�X�v���`���̍זE���Q��p�Ɋ��������悤�ɂȂ����B
�ȏ�̌��ʂ́A�x����̂��זE�W�c�̑��݂������A�V�X�v���`���̍זE���Q���ʂɑ���זE���x����זE�̊������߂鎡�Ð헪�Ƃ��Ă�ALDH1�̑j�Q�̗L�������������Ă���B
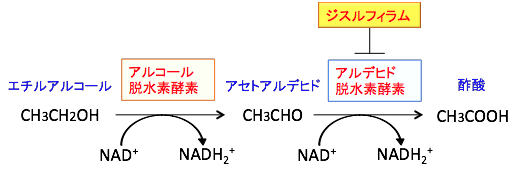
�V�X�v���`���icisplatin : CDDP�j�͔������̂ɕ��ނ����R������ᇍ܁i�R����܁j�ł��B 2�̉��f���q���ʂ�DNA�ƌ������邽�߁ADNA�����ɉˋ����`������ADNA������j�Q���A�זE���Ă��邪��זE����ѐ���זE�����ł����܂��B �V�X�v���`����1978�N�ɕč��ƃJ�i�_�ŏ��F����A���{��1983�N�ɏ��F����Ă��܂��B
�����̌Ō`����̎��ÂɎg�p����A�����ڂ͍����̂ł����A���̍�p�@�����畛��p�������̂������ł��B
�V�X�v���`���Ŏ��Â��Ă���ƁA�قڊm���ɃV�X�v���`���ϐ��̂���זE���o�����A�V�X�v���`���͂���������Ȃ��Ȃ�܂��B���̍R����܃V�X�v���`���ϐ��̃��J�j�Y����j�~����A�V�X�v���`���̍R��ᇌ��ʂ��p���ł��܂��B
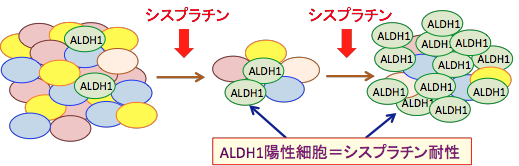
���̘_���ł́A�V�X�v���`��������ёϐ��̔זE���x����זE���ɂ����āA�V�X�v���`���̓��^���A���f�q�h�E���f�y�f�iALDH1�j�̔����̍�������זE�W�c�̏o���܂��͑����U�����A�V�X�v���`���ϐ�������ɍ��߂錋�ʂ�����Ă��܂��B
�܂�AALDH1�����Ă���x����זE�̓V�X�v���`���ɑϐ��̐��������̂ŁAALDH1�z���זE���I��I�ɑ����A�V�X�v���`��������Ɍ����ɂ����Ȃ�Ƃ������ۂ��N���邱�Ƃ��w�E���Ă��܂��B �V�X�v���`�����^���V�X�v���`���ϐ��זE�𑝂₵�A�V�X�v���`���ϐ��זE��ALDH1�̔��������i���Ă����Ƃ������Ƃł��i�}19�j�B
|
�}19�F����g�D�͗l�X�Ȑ���̂���זE����\������Ă���B�V�X�v���`���𓊗^����ƁA�����̂���זE�͎��ł��邪�A�A���f�q�h�E���f�y�f1�iALDH1�j�����Ă��邪��זE�i�}����ALDH1�ƋL�ځj�̓V�X�v���`���ɑϐ��������Đ����c��B����ɃV�X�v���`���𓊗^���Ă��A�A���f�q�h�E���f�y�f�P�iALDH�P�j�̔����̍�������זE�̓V�X�v���`���ɑϐ��������̂ŁA�V�X�v���`�����Âɐ����c���đ��B����B���B��������זE�̓A���f�q�h�E���f�y�f�z������זE�iALDH�P�j�̊����������A����ɃV�X�v���`���ϐ������������B |
����ɁAALDH1�z���זE�̓V�X�v���`���ϐ������łȂ��A�s�ϓ�����⑽�\�����זE�֘A��`�q�̔��������Ȃ����זE�iCancer Stem Cell�j�̐���������ƕ��Ă��܂��B
���זE�̈ێ��ɕK�v�Ȉ�`�q�Ƃ���NANOG, OCT-4, SOX-2, KLF4 , C-MYC�Ȃǂ��m���Ă��܂��B �זE������������iPS�זE����鎞�ɓ�������邢����R�����q�Ƃ����̂́AOct3/4�ASox2�AKlf4�Ac-Myc�̎l�ł��B���̂S�͂��זE�̈ێ��ɂ��K�v�ł��BNanog �͑��\�������艻��������q�ƌ����Ă��܂��B ���̂悤�Ȃ��זE�̐����ێ����K�v�Ȉ�`�q��ALDH1�z���זE�ɑ����������Ă���Ƃ������Ƃł��B
ALDH1�i�A���f�q�h�E���f�y�f�P�j�͂��זE�̃}�[�J�[�ŁA ALDH1��������������זE�͂��זE�̐����������Ă��邱�Ƃ������̌����Ŗ��炩�ɂȂ��Ă��܂��B
���זE�ɓ��ٓI�ȃ}�[�J�[�Ƃ���CD133��ALDH1�Ȃǂ�����܂��B D133�͍זE���ɋǍ݂����100 KDa ��5�ђʌ^�̃^���p�N���ŁA���זE�̓����P���̂��߂̕W�I�\�ʍR���Ƃ��Ďg���Ă��܂��B�������A��e�̂Ȃ̂��A�ڒ����q�Ȃ̂��A���̋@�\�ɂ��Ă͈ˑR�Ƃ��ĕs���ȓ_�������c���Ă��܂��B
ALDH1���A���f�q�h�E���f�y�f�P�ialdehyde dehydrogenase 1�j�ł��B�����ŁA�ȉ��̂悤�ɗ����ł��܂��B
�@ALDH1��������������זE�͂��זE�̐����������Ă���BALDH1�͂��זE�}�[�J�[�Ƃ��ė��p����Ă���B
�A�V�X�v���`���ϐ��̔זE���x����ł�ALDH1�����������BALDH1�����������x����זE�̓V�X�v���`���ɑϐ��ɂȂ�B
�����ŁA ALDH1��j�Q����W�X���t�B�����𓊗^����ƁA�V�X�v���`����R���̔x����זE���V�X�v���`���̍זE���Q��p�Ɋ��������悤�ɂȂ�܂����B
�܂�AALDH1��j�Q����W�X���t�B�����̓V�X�v���`���ϐ���j�~���邱�Ƃ��ł���Ƃ������_�ł��B �W�X���t�B�����ƍR����܂p�����Տ������̌��ʂ�����Ă��܂��B�ȉ��̂悤�ȕ�����܂��B
A phase IIb trial assessing the addition of disulfiram to chemotherapy for the treatment of metastatic non-small cell lung cancer.�i�]�ڐ��זE�x����̎��Â̂��߂̉��w�Ö@�ւ̃W�X���t�B�����̕��p��]�������IIb���Տ������B�j Oncologist. 2015 Apr;20(4):366-7.
��II���Տ������́A������̊��҂�ΏۂɁA�L�����E���S���E���ԂȂǂ̌������s�������ł��B�T���I�ȑO����II���iIIa�j�ƌ��ؓI�Ȍ����II���iIIb�j�ɕ������邱�Ƃ�����A���̘_���̑�IIb���Տ������͗L�����̌����s���āA��III���ɑ�����悤�ȗՏ������ł��B
�זE���x����̍R������Ái�V�X�v���`���{�r�m�����r���j�ɃW�X���t�B�����p���鎡�Â̈��S���ƗL������]�����邽�߂̑�II���Տ��������A���{�݂̃����_������d�ӌ������Ŏ��{���Ă��܂��B
�V�K�ɐf�f���ꂽ�X�e�[�W�S�̔זE���x����40�Ⴊ�R����܁i�V�X�v���`���{�r�m�����r���łQ�T�C�N���ȏ�j�Ŏ��Â��Ă��܂��B����40��̂���20��̓W�X���t�B�����i�P��40mg���P���R��j�p���Ă��܂��B
�������Ԃ̕��ς́A�R����ܒP�ƌQ��7.1�����ŁA�R����܁{�W�X���t�B�������p�Q��10�����ł����B�����������҂��Q�����āA�ǂ�����W�X���t�B�������p�Q�ł����B�W�X���t�B�������p�ɂ����S���ɂ͖��͂���܂���ł����B
�܂�A�X�e�[�W�S�̔זE���x����̃V�X�v���`���{�r�m�����r���̍R������ÂɃW�X���t�B�����p����ƁA�������Ԃ��������ʂ�����Ƃ������ʂł��B
�y�A���f�q�h�E���f�y�f�j�Q�܂̃W�X���t�B�����̓Q���V�^�r���ϐ���j�~����z
�A���f�q�h�E���f�y�f�j�Q�܂̃W�X���t�B�������X������̃Q���V�^�r�����Â̌��ʂ����߂�\��������Ă��܂��B�ȉ��̂悤�ȕ�����܂��B
Aldehyde dehydrogenase 1A1 confers intrinsic and acquired resistance to gemcitabine in human pancreatic adenocarcinoma MIA PaCa-2 cells�i�A���f�q�h�E���f�y�f1A1�̓q�g�X���B����זEMIA PaCa-2�זE�̃W�F���V�^�r���ɑ�����ݐ��ɔ�������ϐ�����ъl�������ϐ��Ɋ֗^����jInt J Oncol. 2012 Sep; 41(3): 855–861.
�y�v�|�z
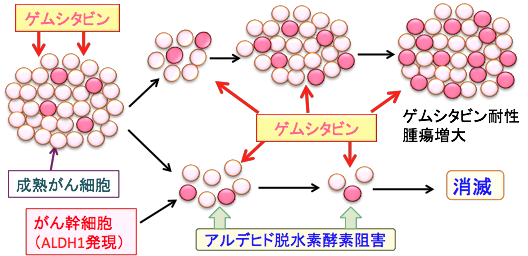
�Q���V�^�r���iGemcitabine�j���X������̍R������ÂɕW���I�Ɏg�p����Ă��邪�A�Q���V�^�r���ɑ���ϐ������Â̗L������W���Ă���B �����̎�ނ̂���ɂ����āA�A���f�q�h�E���f�y�f1A1�̔������x���Ɗ����̘��i�����זE�iCancer stem cells�j�̏d�v�ȓ����Ƃ��Ēm���Ă���B
���זE�̓����̈�Ƃ��čR����܂ɑ���ϐ�������B�{�����ł́A�A���f�q�h�E���f�y�f1A1�iALDH1A1�j�̔����Ɗ����̍����q�g�X������זE���iMIA PaCa-2�זE�j�ɂ������������ALDH1A1�̔������x������ъ����ƃQ���V�^�r���iGEM�j�ɑ����R���̊֘A�ɂ��Č��������B
�ᕪ�q����RNA(small interfering RNA: siRNA)��p����ALDH1A1�̊��������������AGEM�ϐ��ɂ�����ALDH1A1�̊֗^�����������B �ᕪ�q����RNA�ɂ����ALDH1A1�̔����Ɗ����͌����ɒቺ���A�זE���B���j�Q���ꂽ�B ����ɁA�ᕪ�q����RNA�ɂ��ALDH1A1�̃m�b�N�_�E����GEM�̕��p�́A����זE�̐������������ɒቺ�����A�A�|�g�[�V�X�ɂ��זE���𑝂₵�AS���ōזE�������~�����זE�������B
GEN�ϐ��łȂ�MIA PaCa-2�זE���ɔ�ׂāAGEM�ϐ���MIA PaCa-2�זE���ł́AALDH1A1�̔����ʂƊ������L�ӂɘ��i���Ă����B GEM�ϐ���MIA PaCa-2�זE���ɂ����āAALDH1A1�̃m�b�N�_�E����GEM�̕��p�͍זE�̐������������ɒቺ�����A�A�|�g�[�V�X�i�זE���j�𑝂₵���B
���̌��ʂ��A�זE�ɖ{����������ŗL�iintrinsic�j��GEM�ϐ��Ɗl�������iacquired�jGEM�ϐ��̗����ɂ�����ALDH1A1�̊֗^���d�v�ł��邱�Ƃ��������Ă���B
�X�������GEM���Âɂ����āA�ᕪ�q����RNA�ɂ��ALDH1A1�̃m�b�N�_�E����GEM�̕��p�́AGEM�ϐ�����������L���Ȏ��Ö@�Ƃ��āA����Ɍ�����[�߂�K�v������B
�Q���V�^�r���iGemcitabine�j�̓s���~�W����ӂ�j�Q���邱�Ƃɂ���čR�����p�����܂��B GEM�͍זE���ŃQ���V�^�r���R�����_�igemcitabine triphosphate: dFdCTP�j�ɑ�ӂ����DNA�Ɏ�荞�܂��DNA������j�Q���A�זE���i�A�|�g�[�V�X�j��U�����܂��B
�i�s�����X������̍R������ÂŃt�@�[�X�g���C���̍R����܂Ƃ��ĒP�Ƃ��邢�͑��̍R����܂ƕ��p���Ďg�p����Ă��܂��B �������A�X������זE�͓������iintrinsic�j���邢�͊l�����iacquired�j��GEM�ɑϐ��������Ƃ����Ì��ʂ�W���Ă��܂��B ���������āA���̂悤�ȍR����ܑϐ���j�~�ł���A�X������ɑ���R������Â̗L���������߂邱�Ƃ��ł��܂��B
�ϐ��l���̃��J�j�Y���Ƃ��ẮA�̎捞�݂��ӁA�זE����B�Ɋւ����`�q�̕ω��i���ł��ɂ����Ȃ�j�Ȃǂ�����܂��B ����ɁA���זE��GEM�ϐ���L���A�A���f�q�h�E���f�y�f1A1�iALDH1A1�j���������i���Ă���AALDH1A1������j�Q����ƁAGEM�ϐ����j�~�ł��邱�Ƃ�����Ă��܂��B
GEM�͍זE���Ŋ����_�f��𑝂₵�A�|�g�[�V�X�𑣐i���܂��B���������āA�����_�f�����������GEM�ɂ�鑝�B�}�����ʂ͒ቺ���܂��B
GEM�̍R��ᇌ��ʂɊ����_�f��̎Y�����i���֗^���Ă���AALDH1A1�͊����_�f�̎Y�������炷�̂ŁAGEM�̍R��ᇌ��ʂ�ቺ�����邱�ƂɂȂ�܂��B �܂�AALDH1A1�̔������i�́AGEM�ɂ���ĎY������銈���_�f����������邱�Ƃɂ���āAGEM�̍זE�Ő���ቺ������Ƃ������J�j�Y������������Ă��܂��B
|
�}20�F����g�D�ɂ͂��זE���������݂���B���זE�͎��ȕ������Ă��זE���ێ�����Ɠ����ɁA���n����זE���������Ă���g�D���\�����Ă���B�R������Âɑ��āA���n����זE�����ł��₷�����A���זE�͗l�X�ȋ@���Œ�R���������B���זE�������c��A����͍ĔR�E�Ĕ�����B�R������Ái�Q���V�^�r���j���J��Ԃ��ƁA�Q���V�^�r���ɒ�R���̂��זE�������c��A���זE�������邱�Ƃɂ���āA����ɃQ���V�^�r����R�����������A��ᇂ͑��傷��B���זE���ߏ�ɔ������Ă���A���f�q�h�E���f�y�f��j�Q����Ƃ��זE�̓Q���V�^�r���Ŏ��ł��₷���Ȃ�A����g�D�����łł���B |
ALDH�ɂ�17��ނ̃A�C�\�t�H�[��������AALDH1A1�͂��̈�ł��B
ALDH1A1�͍זE���̃A���f�q�h���_�����邱�Ƃɂ���Ď_���X�g���X���y�����܂��B �ᕪ�q����RNA���܂Ȃǂ�ALDH1A1�̔����⊈����j�Q����ƁA����זE�̍R����܊������i���邱�Ƃ�����Ă��܂��B
�X������זE��GEM���݉��Œ����Ԕ|�{�����ALDH1A1�z�����X�����זE���U������Ă��邱�Ƃ�����Ă��܂��B �Q���V�^�r���ϐ������łȂ��A�������܂�^�L�T���n�̍R����܂ɑϐ��̗�������A���ܑϐ��݂̈���זE�A�V�N���t�H�X�t�@�~�h�ϐ��̃q�g����זE�Ȃǂ�ALDH1A1�̔������i������Ă���AALDH1A1�͑����̎�ނ̂���זE�ɂ������ܑϐ��̌����ƂȂ��Ă���\������������Ă��܂��B
ALDH1A1�́A������A�咰����A�X������A�x����A�̑�����A��������ȂǑ����̎�ނ̂���ɂ����āA���זE�̃}�[�J�[�Ƃ��ĔF�߂��Ă���A���זE�ɂ����Ĕ��������i���Ă��܂��B�����āA�A�|�g�[�V�X��R����R����ܑϐ��̌����ƂȂ��Ă��܂��B
ALDH1�̔�������������͗\��s�ǂł��邱�Ƃ������̂����ŕ���Ă��܂��B �X�������łȂ��A�R������È�ʂɂ����āAALDH1A1�̑j�Q�́A�R��ᇌ��ʂ����߂���ʂ����҂ł��܂��B���������āA�R������Â�ALDH1A1�̑j�Q�܂ł���W�X���t�B�����̕��p�͎����Ă݂鉿�l������܂��B
�y�W�X���t�B�����̓A���f�q�h�E���f�y�f��j�Q����z
�A���f�q�h�E���f�y�f�iALDH�j�̓A���f�q�h�iCHO�j���_�����ăJ���{���_�iCOOH�j�ɂ��锽����G�}����y�f�ł��B�A���f�q�g�E���f�y�f�͂��זE�̃}�[�J�[�Ƃ��Ă��m���Ă��܂��B�܂�A�A���f�q�g�E���f�y�f�͂��זE�ɉߏ�ɔ������A���̐�����B�⎩�ȕ����ɉ��炩�̏d�v�ȓ������s���Ă��邱�Ƃ��w�E����Ă��܂��B
�זE�ɂƂ��ăA���f�q�h�͓Ő�������̂ŁA�A���f�q�h�𑁂���ӂ��邽�߂ɕK�v�Ȃ̂ł��B �����̂����ɂ�����ALDH�����̍�������זE�͑��B��]�ڂ𑣐i���邱�Ƃ�����Ă��܂��B
ALDH�����̍�����ᇂ͐������Ԃ��Z���\�オ�������Ƃ�����Ă��܂��B ALDH������j�Q����Ƃ���זE�̑��B��]�ڂ�}���ł��A�R����܂̌����ڂ����߂邱�Ƃ��ł��܂��B
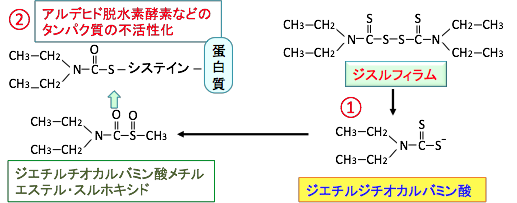
�W�X���t�B�����iDisulfiram�j�̓A���f�q�h�E���f�y�f��j�Q�����p������A�f���Ƃ��ăA���R�[�����ł̎��Ö�Ƃ���60�N�ȏ�O���珈����Ƃ��Ďg�p����Ă��܂��B�A���R�[�������ނƋ�������p���o�܂����A�A���R�[���������܂Ȃ���A�W�X���t�B�����͕���p�̋ɂ߂ď��Ȃ���ł��B �A���f�q�h�E���f�y�f�̑j�Q�܂ł���W�X���t�B�����̓A���R�[���̑�ӂłł���A�Z�g�A���f�q�h�̕�����j�Q���邱�Ƃɂ���āA�A�Z�g�A���f�q�h�̗L�Q�ȏǏł�̂ŁA�A���R�[�������߂Ȃ�����̂ł��B
|
�}21�F�G�`���A���R�[���i�G�^�m�[���j�̓A���f�q�h�E���f�y�f�ŃA�Z�g�A���f�q�h�ɑ�ӂ���A�A�Z�g���f�q�h�̓A���f�q�h�E���f�y�f�ɂ���Đ|�_�ɑ�ӂ����B�W�X���t�B�����̓A���f�q�h�E���f�y�f��j�Q����B�A�Z�g�A���f�q�h�͓Ő��������̂ŁA�זE��g�D�Ƀ_���[�W��^����B |
�W�X���t�B������ALDH1A1��j�Q���܂��BALDH1A1�����łȂ��A����ALDH�A�C�\�t�H�[����s�t�I�ɑj�Q�����ALDH�j�Q�܁ipan-ALDH inhibitor�j�ł���A�זE������у~�g�R���h���A��ALDH���L�͂ɑj�Q���܂��B
���̑��A�W�X���t�B�������v���e�A�\�[���iproteasome�j�ɂ�����^���p�N���̕����@�\�����͂ɑj�Q�����p������܂��B�_���X�g���X�����߂��p������܂��B
�A���R�[����ێ悵�Ȃ���A�ʏ�̕��p�ʂł̓W�X���t�B�������̂̓Ő��͋ɂ߂Čy���ł��B�R������ÂƂ̕��p���A����p�������p�͔F�߂��Ă��܂���B ���������āA�R������Âƕ��p���čR��ᇌ��ʂ����߂���@�Ƃ��āA�W�X���t�B�����͋ɂ߂ėL�p�ł��B�i�������A�p�N���^�L�Z���̂悤�ɗn���ɃG�^�m�[�����g���ꍇ�͎g�p�ł��܂����j
|
�}22�F�W�X���t�B�������o���ێ悷��ƁA�����Ǔ�����ь��t���łP���q�̃W�X���t�B�����͂Q���q�̃W�G�`���W�`�I�J���o�~���_�ɕϊ�����A����ɃW�G�`���`�I�J���o�~���_���`���G�X�e���E�X���z�L�V�h�ɑ�ӂ����i�@�j�B�W�G�`���`�I�J���o�~���_���`���G�X�e���E�X���z�L�V�h�́A�A���f�q�h�E���f�y�f�̍y�f�������ʂ̃V�X�e�C���̃X���t�q�h������i-SH�j�Ɣ������Č������A�y�f������j�Q����i�A�j�B���l�ȃ��J�j�Y���i�^���p�N���̃V�X�e�C���ɔ������Ċ�����j�Q����@���j�ɂ���āA�v���e�C���L�i�[�[C��P���`������DNA���`���g�����X�t�F���[�[���܂ށA�l�X�Ȃ��i���̃^���p�N����j�Q����B |
�����ɑ����܂܂���h�R�T�w�L�T���G���_�iDHA�j�ƃW�X���t�B�����iDSF�j�p����ƁA����זE�̎_���X�g���X�����i���A���זE�̐����iStemness�j���ቺ���čR����܊������i����Ƃ������ʂ�����Ă��܂��B
�R������Ò���ω�R�n�in-3�n�j�����s�O�a���b�_��DHA��EPA��ێ悷��ƁA�R����܂̕���p���y�����A�t����������Ȃǂ̍R��ᇌ��ʂ����߂邱�Ƃ��ł��邱�Ƃ͑����̗Տ������Ŏ�����Ă��܂��B
�h�R�T�w�L�T�G���_�iDHA�j�͐���זE�ɂ͈��e����^�����ɁA����זE�̎_���X�g���X�����߂邱�Ƃ��m���Ă��܂��B DHA�͂���זE�ɑ��Ď_���X�g���X�����邱�Ƃɂ���Ă���זE�̃A�|�g�[�V�X��U�����A�R����܊������߂�Ƃ������ʂ�����Ă��܂��B �W�X���t�B�������_���X�g���X�����߂܂��B �W�X���t�B�������x����̍R������Â̌����ڂ����߂�Ƃ��͑O�q���܂����B
�R������Ò��ɃW�X���t�B�����Ƌ����i���̂̂������j���邢�̓h�R�T�w�L�T�G���_�iDHA�j�̓������T�v�������g�p����̂́A�����Ă݂鉿�l������Ǝv���܂��B
�W�X���t�B�����̍R�����p�͂��Ȃ�Â��i1970�N�ケ��j���猤������Ă��܂����A2010�N�ォ�炻�̍R�����p�����Ȃ蒍�ڂ����悤�ɂȂ��Ă��܂��B �W�X���t�B�����̒f����p�̓A���f�q�h�E���f�y�f�̑j�Q�ɂ��̂ł����A�A���f�q�h�E���f�y�f�͂��זE�̃}�[�J�[�ɂȂ�قǁA���זE�ł̓A���f�q�h�E���f�y�f�͏d�v�ȓ��������Ă���A���̍y�f��j�Q����ƁA����זE�̍R����܊������i���邱�Ƃ����炩�ɂȂ��Ă��܂��B
�W�X���t�B�����͂���̑�֗Ö@�̗̈�ł͐��N�O���痘�p����Ă���A���̍R�����p�̃��J�j�Y���������̕�����܂��B
����2014�N�����炪�ÂɃW�X���t�B������ϋɓI�ɗ��p���Ă��܂����A�m���ȍR�����p��F�߂Ă��܂��B��r�I�����ł���A�A���R�[���������܂Ȃ���W�X���t�B�����͋ɂ߂ĕ���p�̏��Ȃ���ł��B
�i���F�h�Z�^�L�Z���̂悤�ɗn���ɃG�^�m�[�����g������̂ŁA�_�H���Ă���Ƃ��́A�G�^�m�[���t���[�ł��邱�Ƃ��m�F���邱�Ƃ��d�v�ł��j
�߃����E�}�`�̎��Ö�ł����I�[���m�t�B���̓`�I���h�L�V���Ҍ��y�f��j�Q�����p������A�W�X���t�B������DHA�̍R��ᇌ��ʂ�����ɍ��߂܂��B
�܂�A�R����܂���ː����Â̌����ڂ����߂���@�Ƃ��ăW�X���t�B�����{DHA�{�I�[���m�t�B���͎����Ă݂鉿�l�͂���܂��B���ۂɑ����̊��҂���Ɏg�p���Ă��܂����A����p���قƂ�ǂ���܂���B�R��ᇌ��ʂ��m�F���Ă��܂��B
�y��p�z
���ÂɊւ��邲����₲���k�̓��[���iinfo@f-gtc.or.jp�j���d�b�i03-5550-3552�j�ł��⍇���������B
�ȉ���30�����̔�p�̖ڈ��ł��B�̏d��a��ɂ���ĕ��p�ʂ��������邱�Ƃ�����܂��B���ꂼ��̖�уT�v�������g�̖��O���N���b�N����ƁA����̃T�C�g�Ɉڍs���܂��B
| ���i���邢�̓T�v�������g�� |
�P�����p�� |
30�����̔�p�̖ڈ� |
| 2-�f�I�L�V-D-�O���R�[�X |
20�`60mg/kg�̏d |
20,000�~�`50,000�~ |
| ���g�z���~�� |
500mg�`1500mg |
3,000�~�`9,000�~ |
| �W�N�����|�_�i�g���E�� |
10�`20mg/kg�̏d |
10,000�~�`20,000�~ |
| R����|�_���Z���� |
4�`�W�J�v�Z�� |
5,000�~�`10,000�~ |
| �r�^�~��B�P�i�x���t�H�`�A�~���j |
100�`150mg |
2,700�~�`3,600�~ |
| �W�X���t�B���� |
250mg |
12,000�~ |
| �I�[���m�t�B�� |
6 mg |
6,000�~ |
���F�W�X���t�B�������p���͈����͂ł��܂���B�ޗǒЂ��̂悤�ȃA���R�[���̓������H�i���H�ׂ�܂���B�R����܂̃p�N���^�L�Z���͗n���܂Ƃ��ăG�^�m�[����p���Ă��܂��̂ŁA�p�N���^�L�Z�����Ò��̓W�X���t�B�����͎g�p�ł��܂���B�_�H�ɂ��R������Â��Ă���Ƃ��ɂ́A�n���܂ȂǂŃG�^�m�[�����g�p���Ă��Ȃ����Ƃ��m�F����K�v������܂��B
����ɁA����̃A���J���Ö@�p����ƁA����ɍR��ᇌ��ʂ����߂邱�Ƃ��ł��܂��B
�i�A���J���Ö@�ɂ��Ă͂�������j
�R����܂����ł͌��E������܂��B�ȏ�̕��@�͂P�����̔�p���U�`�W���~���x������܂����A �i�s����ł������ł���\�������߂܂��B
���@�ł͑����̕⊮�E��֗Ö@�����H���Ă��܂��B���ꂼ��̎��Ö@�̍�����L�������������߂ɁA���ꂼ��̃y�[�W�ŏڂ���������Ă��܂��B�������A����̎�ނ�i�s�x�⎡�Â̏ɂ���čœK�Ȏ��Ö@�͈قȂ�܂��B���ꂼ��̎��Ö@�𗝉����A�����ɍ��������Ö@��I�����邱�Ƃ́A���҂���ɂ͍���Ǝv���܂��B ���݂̕a��⎡�Â̏����[���iinfo@f-gtc.or.jp�j���d�b�i03-5550-3552�j���t�@�b�N�X�i03-3541-7577�j�ł��m�点����������A�S�ĉ@���i���c��T�j�����������Ă��܂��B�����@����������A�ڂ����������������܂��B